The testes weight of Spo11βki/- mice varies with the genetic background
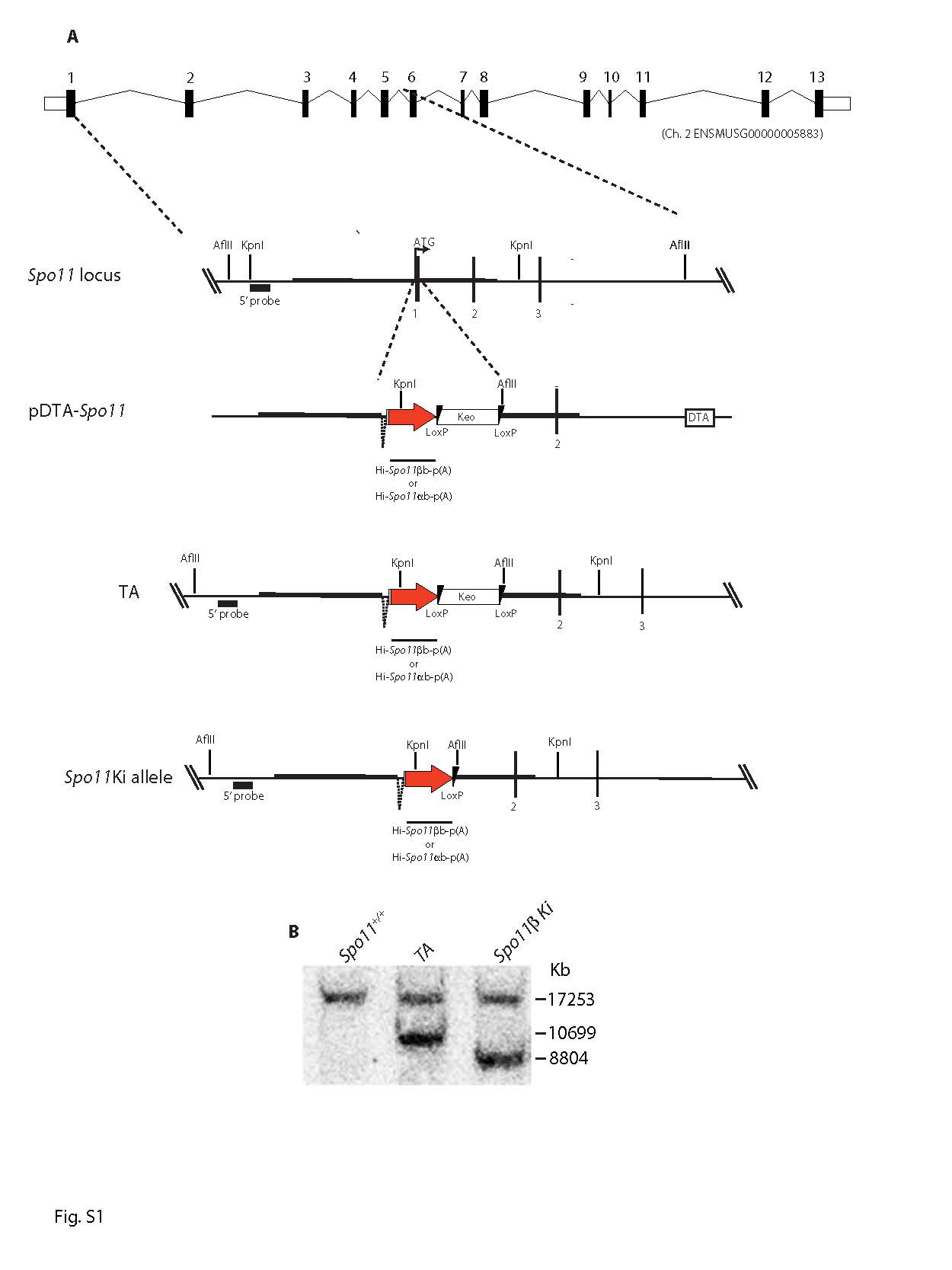
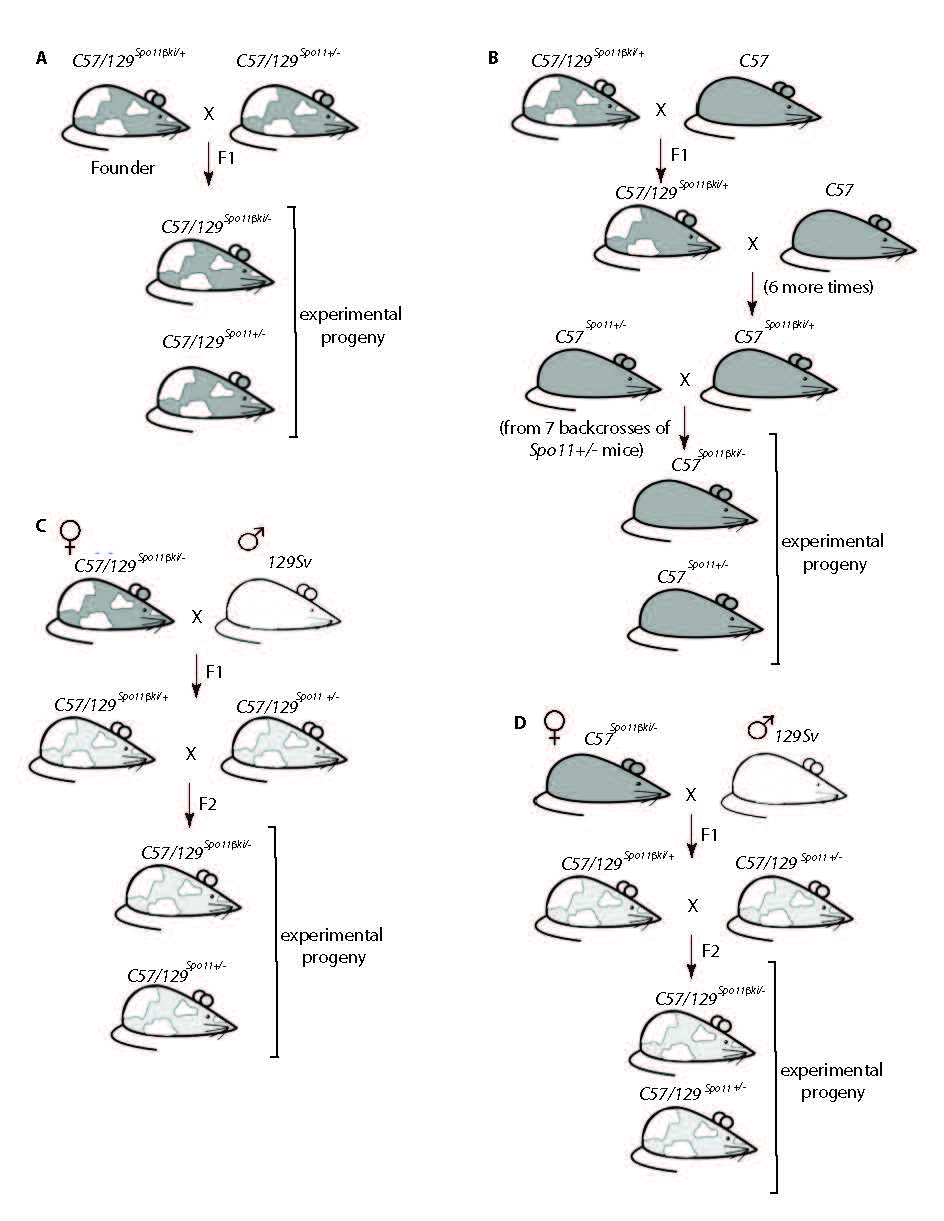
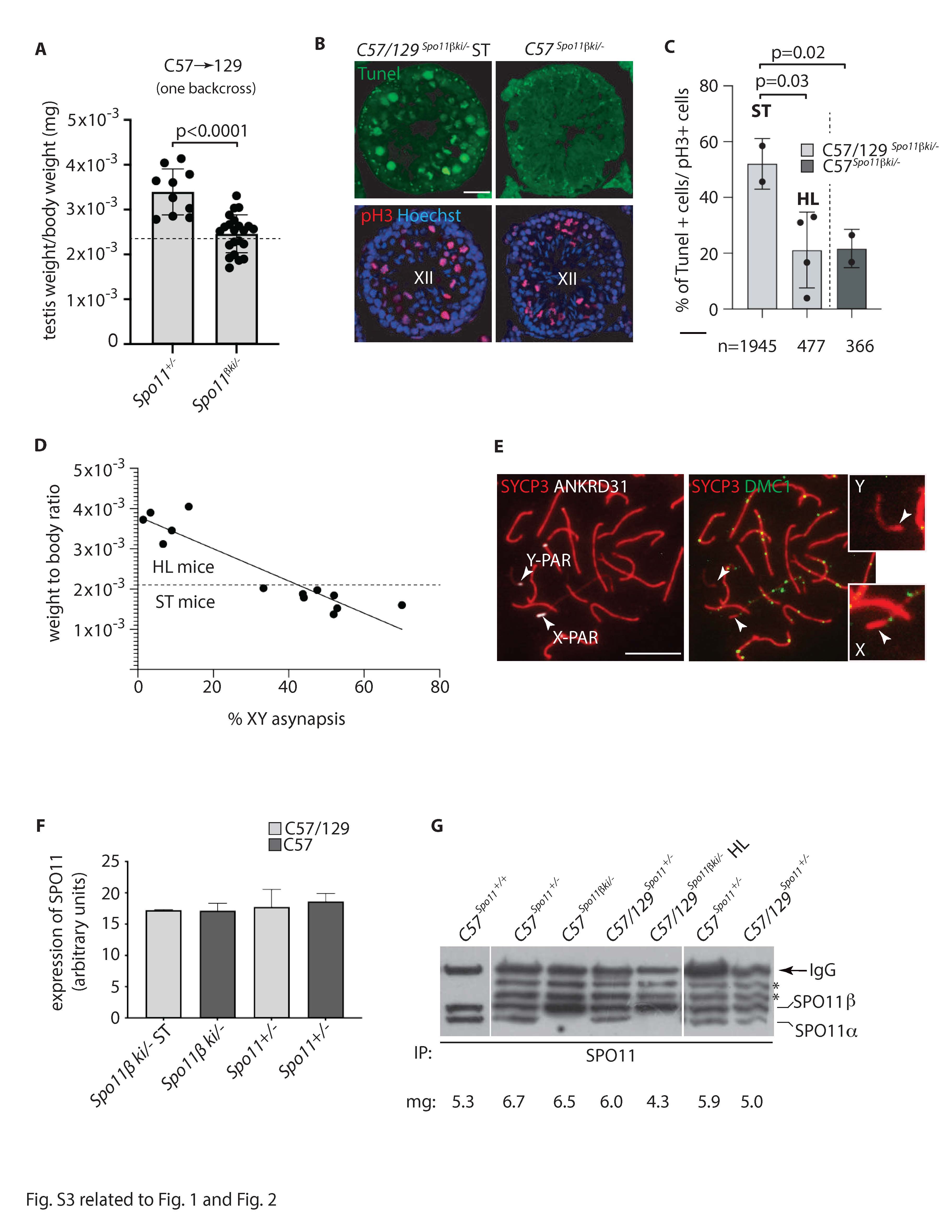
In male mammals, death of defective germ cells within the testis, causes an overall reduction in testis weight, so this can be used to quantify spermatogenesis performance (e.g., see [32]).To test how the expression of Spo11β affects spermatogenesis when the protein is expressed under normal physiological timing and at allelic dosage, we generated mice expressing a single knock-in allele of Spo11βb (thereafter named Spo11βki/-) under the control of the Spo11 promoter (Fig. S1). Mice were created with a mixed (C57BL/6 and 129Sv) genetic background (C57/129Spo11βki/-), see material and methods and Fig. S2A. Examination of relative testis weight (testis to body-weight ratio) revealed great variability among C57/129Spo11βki/- mice, compared to littermates C57/129Spo11+/-. Indeed, while some C57/129Spo11βki/- males had testes with visibly reduced weights, below the mean (i.e., small testis; ST), others appeared indistinguishable from Spo11+/- mice (i.e., with a het-like (HL) phenotype) (Fig. 1A). Nevertheless, relative testis weight of ST mice was greater than in Spo11-/- mice, in which progression of meiosis arrests at zygonema of the first meiotic division [5, 6, 32], indicating that in C57/129Spo11βki/- mice the arrest is either incomplete or it occurs beyond zygonema. Given that the mice were of mixed genetic background, we reasoned that the observed phenotypic variability could have been related to background variations. To test this interpretation, we introduced the Spo11βki allele into a pure C57/BL6 background (C57Spo11βki/- mice) (see Fig. S2B and material and methods); variability was greatly reduced, and testis to body weight ratio turned very similar to Spo11+/- (Fig. 1B). Next, to understand whether the phenotype would have worsened in the 129Sv background, we backcrossed C57/129Spo11βki/- mice into 129Sv for one generation (see Fig. S2C and material and methods). A single backcross shift was sufficient to worsen the phenotype (compare C57/129Spo11βki/- mice in Fig. 1A and Fig. 1C). This was also confirmed by backcrossing C57Spo11βki/- mice into 129Sv for one generation (Fig. S2D and S3A). We concluded that in males with the Spo11βki/- genotype, the performance of spermatogenesis changes with genetic background.
Reduced testis-weight to body ratio in C57/129 Spo11βki/- ST mice correlate with failure of sex chromosome synapsis and apoptotic elimination of spermatocytes at metaphase I
In mammals, synapsis of spermatocyte chromosomes occur in the context of the development of a zipper-like proteinaceous structure called synaptonemal complex (SC) [33]. Synapsis begins with the alignment of the homologs at leptonema and is completed by pachynema. Cytologically, cells in leptonema are identified by the appearance of SYCP3 positive stretches of the lateral elements of the SC; progression to zygonema is marked by the assembly of the SYCP1-positive central element of the SC, between pairs of synapsed homologues. At pachynema, autosomes are fully synapsed throughout their entire length and SYCP3 and SYCP1 signals overlap throughout. In contrast, synapsis between XY chromosomes occurs only at the PAR. Thus, a short stretch of SYCP1 forms between chromosomes, only in this region, while the rest of the chromosomes axes is marked by SYCP3.
To probe if variations in relative testis weight in C57/129Spo11βki/- mice was related to the proficiency of XY synapsis, we quantified XY asynapsis in our genotypes of interest by staining surface spread chromosomes of C57/129 Spo11βki/- ST, C57/129 Spo11βki/- HL and C57Spo11βki/- males with anti-SYCP3 and anti-SYCP1 antibodies. While in C57/129 Spo11βki/- ST, XY synapsis failed in ~ 55% of spermatocytes; the percentage was down to ~ 11% in C57/129 Spo11βki/- HL and to 4% in C57 Spo11βki/- mice (Fig. 1D-E), indicating that the reduced testis weight and frequency of XY asynapsis are closely correlated.
In male mice and humans, each seminiferous tubule cross section can be assigned to one of the 12 epithelial stages (numbered I-XII) based on the array of germ cell developmental stages it contains [34–36]. Elimination of MI spermatocytes that have achiasmate homolog pairs (non-exchange) occurs in stage XII by activating the spindle assembly checkpoint (SAC) [14, 37]. To evaluate the occurrence of germ cell loss by apoptosis at stage XII, we combined terminal deoxynucleotidyl transferase dUTP nick end labelling (TUNEL) and anti-H3Ser10 (pH3) staining in testis sections. The latter was used as a marker to identify metaphase I (MI) cells in stage XII. As shown in Fig. S3B and quantified in Fig. S3C, the frequency of MI cell apoptosis was higher in the tubules of C57/129 Spo11βki/- ST males compared to those of C57/129 Spo11βki/- HL and C57Spo11βki/- males. We concluded that in mice with a Spo11βki/- genotype testicular atrophy is related to failure of XY synapsis and apoptotic elimination of defective spermatocytes in stage XII.
C57/129 Spo11βki/- ST spermatocytes are defective for the formation of DSBs in the PAR
To assess whether the defect of XY synapsis in C57/129 Spo11βki/- ST mice was attributable to the lack of DSBs formation in the PAR, we combined the staining of the SC component SYCP3 and DMC1 (a surrogate marker of DSBs [12, 38, 39]) with that of PAR, using fluorescent in situ hybridization (FISH). The PAR probe recognizes a region at the boundary between the non-PAR region and the PARs of the X and Y chromosomes, and hybridizes with the tandem array of minisatellite mo-2 at the noncentromeric end of chromosomes 4, 9 and 13 [23]. This prevents unequivocal identification. On the contrary, the Y-PAR FISH signal has a distinctive pattern, as the FISH staining always extends from the Y chromosome axis to the chromatin loops, forming a distinguishable cloud around the Y-PAR (Fig. 2A). Under physiological conditions, DSB formation occurs with a comparable frequency in both the X-PAR and Y-PAR, mainly at the late zygotene stage, [14]. Therefore, since the Y-PAR is uniquely identified with the PAR FISH probe, we quantified the frequency of DMC1 foci in this region, in late zygonema spermatocytes from C57/129Spo11βki/- ST and C57Spo11βki/-mice. To enrich our samples for germ cells at late zygonema, we prepared chromosome spreads from 12 dpp mice. At this time point, apoptosis selection of cells defective in XY synapsis had not yet occurred [32], therefore, the ST phenotype cannot be assessed. To overcome this problem, we evaluated the percentage of XY asynapsis and only included C57/129Spo11βki/- mice with at least 35% XY asynapsis in the analysis (Fig. 2B). This value was set according to the correlation between the frequency of XY asynapsis and testis-weight to body ratio in adult C57/129Spo11βki/- mice (Fig. S3D). Alongside, with this, we analyzed DSBs formation in C57Spo11βki/- spermatocytes, in which the average XY asynapsis was less than 10% (Fig. 2B). The analysis of the presence of DMC1 foci in the PAR of late zygotene cells, revealed that the high degree of XY asynapsis correlates with a reduced frequency of the presence of DMC1 foci (Fig. 2C). However, the frequency of DMC1 in the Y-PAR was low compared to the percentage of XY asynapsis. This raised the question of whether DSBs form more frequently in the X-PAR than in the Y-PAR. To test this, we identified both PARs by immunolocalizing ANKRD31, which at the zygotene-pachytene transition and at the early pachytene stages aggregate on PARs (see [26] and below). We analyzed spermatocytes of C57/129 Spo11βki/- 14 dpp mice with an average XY asynapsis (estimated by SYCP3/SYCP1 staining) equal to 52.5± 8%. Of 42 cells with unsynapsed sex chromosomes, 25 (59,5%) had no foci on PARs (Fig. S3E), 10 (23,8%) showed a focus only on the X-PAR, 5 (12%) only in Y-PAR, and 2 (4,7%) in both PARs. The latter are likely cells in which foci are found on both chromosomes, upon release of one DSB from either PARs [14]. We concluded that in C57/129 Spo11βki/- spermatocytes XY asynapsis occurs as a result of the lack/delayed formation of DSBs on PARs, confirming previous findings [14], and that the frequency of DSBs in the Y-PAR is about twice as low as in the X-PAR.
Analysis of SPO11 expression in mice with different genetic backgrounds
In mice, the expression of the SPO11 protein below a critical amount may have an effect on DSB levels and chromosome synapsis proficiency [38, 39]. To test whether failure of XY synapsis in C57/129 Spo11βki/- ST mice was related to faulty expression of SPO11β, we immunoprecipitated it from mouse testis extracts from juvenile mice at 12 dpp. Protein levels among Spo11+/- and Spo11βki/- mice were comparable (Fig. 2D and Fig. S3F). SPO11 protein levels were also comparable among C57/129Spo11+/-, C57Spo11βki/- and C57/129 Spo11βki/- HL genotypes, in adults (Fig. S3G). Next, to investigate whether SPO11 function is normal in C57/129 Spo11βki/- ST spermatocytes, we quantified the number of DSBs nucleus wide by co-staining spermatocyte surface chromosome spreads with SYCP3 and DMC1. We did not observe a reduction in DMC1 foci number in C57/129Spo11βki/- ST cells compared to C57Spo11βki/- spermatocytes. Rather, the average number of foci at leptonema and early mid-zygonema increased slightly in C57/129 Spo11βki/- ST cells (Fig. 2E). We concluded that it is unlikely that the reduced frequency of DSB formation in the PAR of C57/129Spo11βki/- ST mice is due to defects of SPO11β expression or function.
Reduced DSB formation in the PAR of C57/129 Spo11βki/- ST spermatocytes is not related to defects in the aggregation of the auxiliary proteins of SPO11.
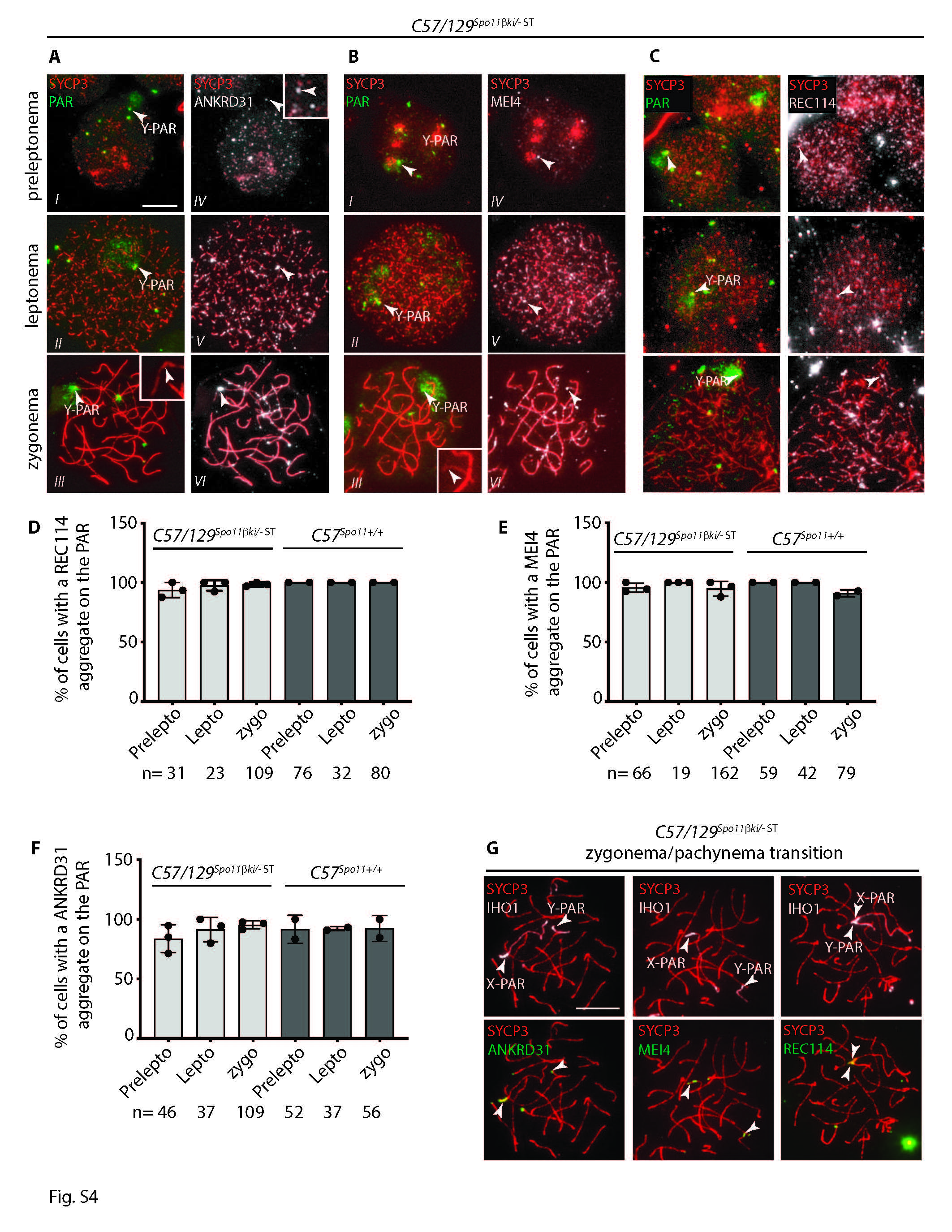
The formation of DSBs in the PAR by SPO11 occurs with the assistance of auxiliary proteins, including IHO1, MEI4, REC114, MEI1 and ANKRD31 (RMMAI complex) [19–26]. Aggregation of RMMAI proteins on the PAR occurs from the preleptotene stage, in advance of the formation of DSBs [23]. To investigate whether SPO11 auxiliary proteins localize normally in PARs of mice with increased XY asynapsis, we monitored the assembly of ANKRD31, MEI4, and REC114 from preleptonema to zygonema in C57/129 Spo11βki/- ST mice. Spermatocytes from wild type C57 mice were used as a control. To identify their association with the PAR axis, surface chromosome spreads were stained with SYCP3 and the PAR probe. As shown in Fig. S4A-C and quantified in Fig. S4D-F, aggregation of these factors was comparable to that of the control. Furthermore, we immunolocalized aggregates of ANKRD31, MEI4 and REC114 at the zygotene/pachytene transition stage, the sub-stage when most DSBs form in the PAR [14]. To this end, we colocalized them with IHO1, which at this stage forms a blob signal only on X-PAR and Y-PAR [20]. In this case, we never observed cells without ANKRD31, MEI4, or REC114 aggregates in C57/129 Spo11βki/- ST mice (197, 168 and 231 cells analyzed respectively, from three mice per genotype) (Fig. S4G). From these results, we ruled out the possibility that a defective aggregation of RMMAI proteins is responsible for the XY asynapsis defects observed in C57/129 Spo11βki/- ST spermatocytes.
Spermatocytes from C57/129 Spo11βki/- ST and C57 Spo11βki/- mice differ in the high-order chromatin structure of the PAR
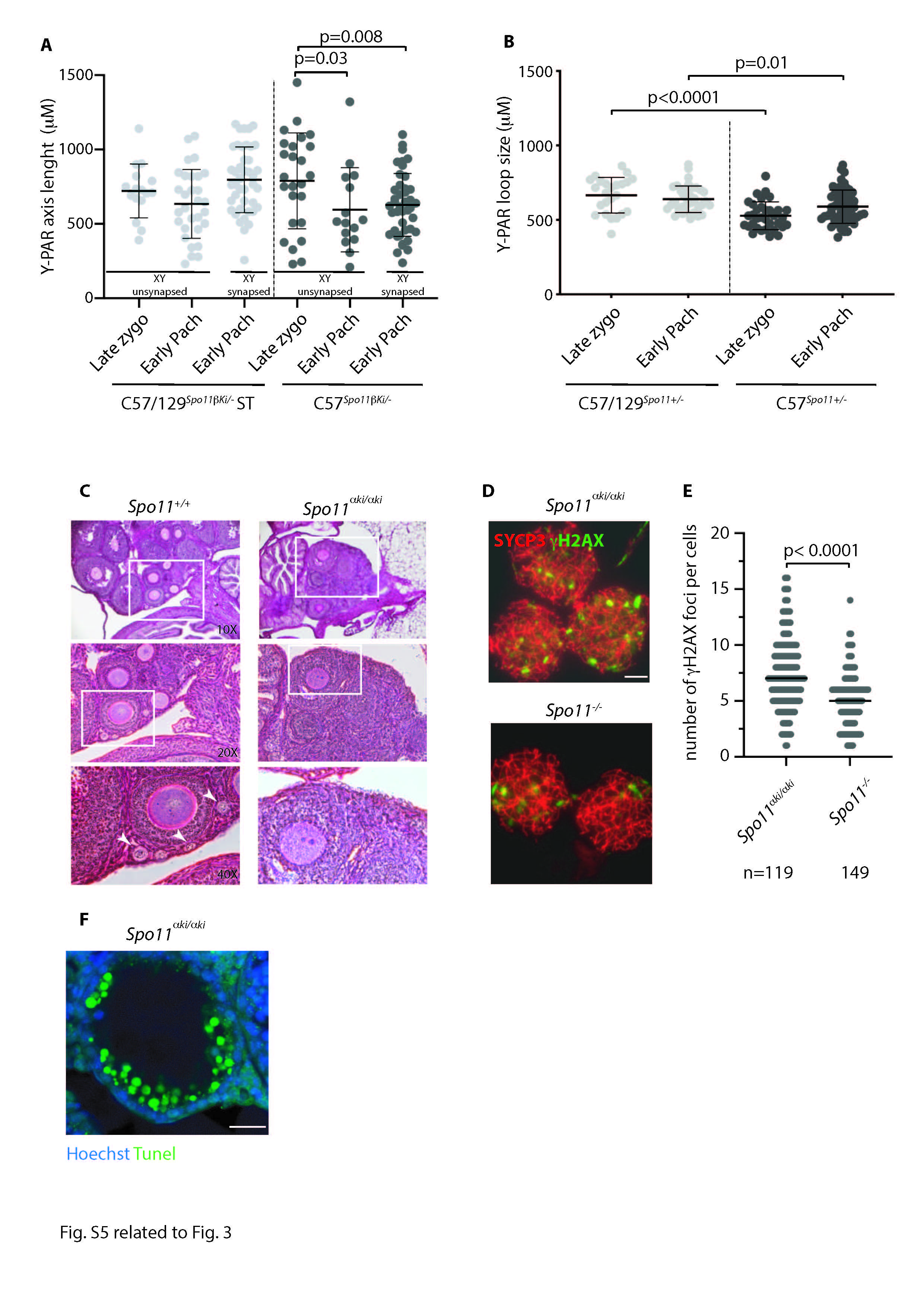
In mice, the formation of DSBs in the PAR is preceded by its ultrastructural remodeling that consists of the separation (splitting) by zygonema of the aligned sister chromatid axes, decorated with RMMAI proteins [23]. To monitor potential changes in PAR ultrastructure in C57/129 Spo11βki/- ST spermatocytes, we analyzed the PAR of surface chromosome spreads of spermatocytes at the zygonema/pachynema transition using Stimulated Emission Depletion (STED) microscopy. To this end, the spermatocyte chromosome axis was stained with anti-SYCP3 antibody, while the sex chromosomes and PARs were identified by IHO1 stain [20]. PARs were also identified by using the anti-ANKRD31 antibody, which forms distinguishable large aggregates on both the X-PAR and Y-PAR [26] (Fig. 3A). By comparing STED images (insets in Fig. 3A), we found that the frequencies of X-PAR axis splitting in late zygonema were comparable between C57/129 Spo11βki/- ST and C57 Spo11βki/- mice we used as control (92%, n = 34 and 91%, n = 33, respectively), while Y-PAR splitting occurred less frequently in C57/129 Spo11βki/- ST mice (C57/129 Spo11βki/- ST 84%, n = 35; C57 Spo11βki/- 96%, n = 41, p = 0.0004 Chi-Square test). Although the physiological role of PAR splitting is still unclear [23], this result suggested a small but noticeable defect in Y-PAR remodeling. In mouse splitting of the PARs axes are strictly temporally correlated with the remodeling of the PAR chromatin loops and axis. The PARs loops are short at leptonema up to late zygonema, when DSBs are made in the PAR, and lengthen in early to mid-pachynema cells [23]. Correspondingly, the PAR axis is long as soon as it is detectable at leptonema and late zygonema/early pachynema and shortens in the mid-pachytene stage [23]. In our effort to understand the molecular basis of the defect of XY synapsis in C57/129 Spo11βki/- ST spermatocytes, we sought to study the changes in PAR conformation by measuring the length of the loops and the axis during prophase I in surface spreads of spermatocytes stained with SYCP3 and the PAR FISH probe. We focused on the Y-PAR, as it is uniquely identifiable and its dynamic changes in wild- type cells are well characterized [23]. As a control, we employed C57Spo11βki/- males, which are more proficient in XY synapsis (Fig. 2C). The size of loops was defined as the axis-orthogonal extension of the PAR FISH signal, while the length of the PAR axis was determined as the distance from the PAR probe to the end of the SYCP3 axis (Fig. 3B-C) [14, 23, 40]. Comparing cells at late zygonema and early pachynema in C57/129 Spo11βki/- ST spermatocytes, the average size of PAR loops at late zygonema was shorter than in early pachynema, confirming previous results [23]. This was true regardless of whether the XY synapses had just occurred at early pachynema (Fig. 3D). Similarly, the Y-PAR loops of C57Spo11βki/- spermatocytes at late zygonema were shorter compared to cells at early-pachynema with synapsed sex chromosomes. An upward trend in average loops length was also observed in early pachytene-stage cells with asynapsed XY, although the difference did not reach statistical significance. Remarkably, the comparison of FISH signals among cells of C57/129 Spo11βki/- ST and C57Spo11βki/- mice indicated that the PAR loops of C57 Spo11βki/- mice were constitutively more compact than those of C57/129Spo11βki/- ST cells (Fig. 3D), consistent with smaller loops. Side-by-side analysis of the length of the Y-PAR axis showed that it shortened slightly in early pachynema cells of C57Spo11βki/- mice, while no significant variations were found in C57/129 Spo11βki/- ST cells (Fig. S5A). The latter was expected, as the shortening of the PAR axis is generally measurable by mid-pachynema [23]. We did not find mid-pachytene cells at the 12 dpp time point; therefore, shortening of the PAR axis at this more advanced stage could not be tested. From these experiments, we concluded that the spermatocytes of C57/129 Spo11βki/- ST and C57 Spo11βki/- mice differ for the high-order chromatin structure of the PAR.
Interplay between PAR ultrastructure and expression of the Spo11 wild type allele
In mice carrying a wild type allele of Spo11 in the mixed background (C57/129Spo11+/-), relative weight of the testes is high and less variable compared with that of C57/129Spo11βki/- mice (Figs. 1A, 1C and S3A). To investigate how these phenotypes correlate with the frequency of XY asynapsis, we quantified it in the genetic models of our interest. As shown in Fig. 3E, sex chromosome asynapsis was less frequent in C57/129 Spo11+/- mice compared to C57/129Spo11βki/- males. This indicates that the expression of the full set of Spo11 splice-isoforms by the wild type allele promotes XY recombination and synapsis better than the Spo11βki allele. The subsequent comparison of XY asynapsis in C57/129 Spo11+/- and C57 Spo11+/- males pointed out that the latter are the most proficient. To test whether this correlated with a shortening of the PAR loops length, we measured it in juvenile C57/129Spo11+/- and C57 Spo11+/- mice. PAR loop length in spermatocytes with a C57 background were significantly shorter (Fig. S5B), confirming our previous results (Fig. 3D). Shortening of PAR loops also correlated with a recovery of XY asynapsis in cells from C57Spo11βki/- males (Fig. 3E). We concluded that reduced length of the PAR loops in the C57 background and the expression of a wild type set of Spo11 splice-isoforms, both impacts on XY recombination, likely by distinct mechanisms, in cooperation with each other.
The function of Spo11β on PAR is boosted by the concomitant expression of Spo11α
SPO11α conserves the catalytically active tyrosine residue of Spo11 required for its DSB formation activity [15, 16]; therefore, it is a potentially catalytically active isoform. With the goal of testing the ability of this isoform to form DSBs, we generated a knock-in mouse model that expresses it under the control of the Spo11 promoter (Fig. S1). Mice homozygous for the Spo11αki allele were generated on a C57 background (C57Spo11αki/αki). Analysis of the morphology and relative testicular weight of these mice revealed that they phenocopied Spo11-/- mice [5, 6] (Fig. 4A-B). Furthermore, histological observation of the ovaries of adult mice revealed that females were also phenotypically similar to Spo11-/- [5, 6], as primordial follicles could not be observed in the cortex (Fig. S5C). Consistent with these observations, staining of spermatocyte spread chromosomes with SYCP3 and SYCP1 antibodies, revealed that, just as Spo11-/- spermatocytes [5, 6], C57Spo11αki/αki cells were not able to progress beyond a zygotene-like stage (Fig. 4C). Successive quantification of the number of DSBs in spermatocytes using DMC1 as a surrogate marker, showed that the number of DSBs was extremely low in C57Spo11αki/αki cells compared to wild type mice, although slightly higher than in Spo11-/- spermatocytes (Fig. 4D-E). To confirm this result, we also quantified the number of γH2AX patches, which mark DSB sites regardless of the DMC1 assembly [41]. Again, numbers of γH2AX patches were slightly increased compared to Spo11-/- mice (Fig. S5D-E). Confirming the failure of proper formation of DSBs, the histological analyses of C57Spo11αki/αki testes revealed that, as previously demonstrated in Spo11-/- mice [5, 32], spermatocytes underwent massive cell death (Fig. S5F). Next, we went one step further by testing whether one of the few DSBs that form in C57Spo11αki/αki spermatocytes occur in the PAR. To this end, we immunolocalized DMC1 in the PAR of surface chromosome spreads of C57Spo11αki/αki cells in combination with SYCP3 and the PAR FISH probe (Fig. 4F). Of the three mice analyzed, we never observed DMC1 foci in the PAR of cells in leptonema (n = 53) and found foci in 6/376 nuclei in the zygonema-like stage (1.2% ± 0.3). Conversely, DMC1 foci were never found in the PAR of Spo11-/- cells at any stage (n = 218 cells, from three mice). We concluded that in C57Spo11αki/αki males, DSBs form with extremely low efficiency on both non-sex and sex chromosomes. To investigate whether such a phenotype was traceable to a low level of the protein, we immunoprecipitated SPO11 from C57 wild type, C57Spo11+/- and C57Spo11αki/αki testes. Samples were collected from 12 dpp mice to compare testes with similar progression of meiosis. SPO11α expression in C57Spo11αki/αki mice was visibly reduced compared to SPO11β in wild type and C57Spo11+/- spermatocytes (Fig. 5A). This suggests that the low frequency of DMC1 foci in C57Spo11αki/αki spermatocytes is at least in part attributable to the low protein level.
Considering that under physiological conditions, SPO11α is expressed in prophase I, later than SPO11β, [6, 15, 42, 43], we speculated that another reason why the proficiency of DSB formation in the PAR and autosomes of C57Spo11αki/αki spermatocytes is low is because it lacks SPO11β. As shown in Fig. 5A, in our Spo11α knock-in model, the protein is expressed with an early timing compared to wild type, as it is already well detected in testes of 12 dpp mice, when in wild type mice is only observed SPO11β. Taking advantage of this characteristic, we generated mice expressing one wild type allele of Spo11 in combination with the Spo11αki allele (i.e., C57Spo11αki/+ mice). After verifying the expression of both splice isoforms (Fig. 5A, right panel), we quantified the number of DSBs in the PAR, comparing it with C57 wild type and C57Spo11+/- spermatocytes, which by this age only express SPO11β. Our prediction was that if the function of SPO11β in the PAR is enhanced by concomitant expression of SPO11α, DSBs should form with greater efficiency in the PAR of C57Spo11αki/+ cells at leptonema and early zygonema compared to cells from control genotypes. This expectation was met. Quantification of DMC1 foci in the Y-PAR of leptotene stage cells revealed that the frequency of DSBs was increased by five folds in C57Spo11αki/+ spermatocytes compared to wild type C57 cells and by over 16 folds compared to cells from C57Spo11+/- mice. A smaller increase was also observed in the early/mid zygotene and early pachytene stages, compared to C57Spo11+/- cells (3.9 and 1.1, respectively) (Fig. 5B-C). From this observation, we concluded that SPO11β function in the PAR is augmented by the concomitant expression of SPO11α. Interestingly, quantification of DMC1 foci on whole chromatin of C57Spo11kiα/+ spermatocytes at leptonema and early/mid zygonema revealed that “global” DSBs increased less (1.4 and 1.1 folds, respectively) than in the PAR (Fig. 5D). This indicates that the expression of SPO11α is mainly functionally related to recombination initiation in the PAR.
C57/129 Spo11βki/- ST mice are prone to sex chromosome aneuploidy in sperm
Previous studies have shown that in male mice prone to sex chromosome asynapsis, fertility and differentiation of aneuploid sperm are functions of the degree of XY asynapsis [44]. If XY pairing fails in not more than ∼50% of sperm, activation of the spindle assembly checkpoint (SAC) does not have an obvious impact on sperm production and mice are fertile [44]. Consistent with the fact that in C57/129Spo11βki/− ST mice XY synapsis fails in ~ 55% of cells, mice had reduced but still abundant spermatozoa in the cauda of the epididymis (Fig. S6A) and were fertile (Table S1). Next, to understand whether such mice generated sperm aneuploid for the sex chromosomes, we subjected cells collected from the cauda of the epididymis to FISH with probes against the X and Y chromosomes. A fluorescent in situ hybridization probe for chromosome 8 served as an internal control for correct identification of aneuploid sperm vs diploid ones (Fig. S6B). In C57/129Spo11βki/− ST mice, the percentage of sperm nuclei containing both X and Y or no sex chromosomes was increased (Fig. S6C). We conclude that C57/129Spo11βki/− ST males are prone to formation of aneuploid gametes.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}