In silico characterization of antisense oligonucleotides (ASOs) targeting L1- MET
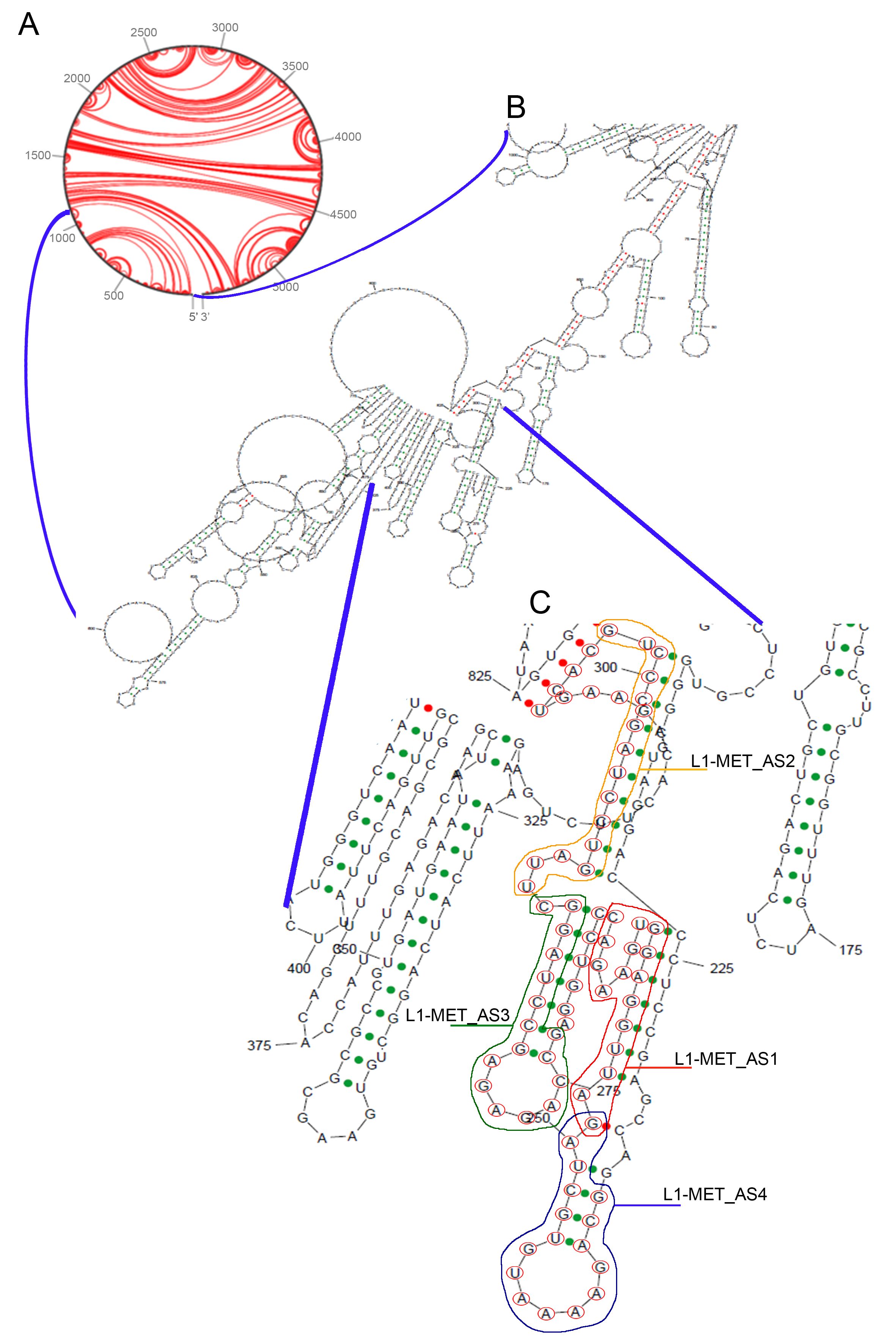
To study the L1-MET expression, we employed modified antisense oligonucleotides (ASOs) able to target specifically L1-MET transcript (Fig. 1A). We identified a specific 76 bp fragment, spanning nucleotides + 176 and + 251 in L1-MET, as the most suitable sequence. The following in- silico analysis allowed to pinpoint 4 out of 11 potential ASOs: L1MET_AS1, complementary to the region between nucleotide + 191 and nucleotide + 206, L1MET_AS2, between + 229 and + 245, L1MET_AS3, between + 213 and + 228, and L1MET_AS4, targeting the beginning of the sequence (from + 176 to + 191) (Table 1 and Fig. 1B).

The ASOs efficiency was then evaluated considering other different parameters. Initially, we used the sRNA software sFOLD to predict their secondary structure and define their level of thermostability. In our hands, three ASOs (L1MET_AS1/2/4) displayed a similar low folding while L1MET_AS3 had a clear hairpin structure (Fig. 1C). The ΔG levels of the four selected ASOs confirmed the L1MET_AS3 as the less stable. Based on these analyses, L1MET_AS1 turned out to be the best ASOs (ΔG = 2.5), whereas L1MET_AS2/4 showed an intermediate value (ΔG = 0.6 and 0.0, respectively). Next, we examined the sequence composition considering T stretches and palindrome sequences detrimental for the ASO activity. We noticed that L1MET_AS4 displayed a 4T stretch in the hairpin loop, strongly impairing the potential effectiveness of the ASO. Finally, the heteroduplex formed by the ASO and the target mRNA was evaluated checking the folding of the entire L1-MET sequence using the sRNA algorithm in sFOLD. Overall, L1MET_AS1 and AS2 targeted the most favorable regions. Due to the poor sequence features and the thermal stability, we excluded L1MET_AS4 from the study, whereas L1MET_AS3 was included, although the predicted stability was poor (Supplementary Fig. 1).
No potential off-target transcripts of the selected ASOs were identified by investigating the transcriptome with NCBI blastn (https://blast.ncbi.nlm.nih.gov/Blast.cgi) and ENSEMBL blastn (https://www.ensembl.org/Homo_sapiens/Tools/Blast). This result was further confirmed even when one mismatch nucleotide was considered.
Corresponding modified Gapmers (LNA-GapmeRs) of the selected ASOs were then developed, as reported in Materials & Methods Section, for the following analyses.
L1-MET expression and silencing
To investigate the biological function of L1-MET, we selected cell lines that expressed variable levels of L1-MET and MET mRNA testing a panel including lung cancer (EBC1 and A549) and breast cancer (MDA-MB231 and MCF7) cells, along with human fibroblasts (MRC5) and normal lymphocytes used as controls.
EBC1 cells showed the highest level of both L1-MET and MET, followed by A549 cells with an intermediate level of L1-MET, but a low level of MET. As for the breast cancer cells, MDA-MB231 weakly expressed both L1-MET and MET mRNA, while MCF-7 lack a detectable MET, but evidenced an intermediate level of L1-MET transcript (Fig. 2A-B).
To study the effects of altered L1-MET expression, the cells were transiently transfected using LNA-GapmeRs corresponding to the ASOs previously selected (L1MET_AS1, L1MET_AS2, and L1MET_AS3). The efficacy of the L1-MET silencing was in line with the in silico prediction and, as shown by qRT-PCR results, a decreasing silencing effect of the three LNA-GapmeRs was observed (Fig. 3). Overall, L1MET_AS2 was the most effective in downregulating the L1-MET transcript in all the cells (EBC1 p < 0.0001; A549 p = 0.0039; MDA-MB231 = 0.0236; MCF7 p = 0.0159). A significant downregulation was also detected using L1MET_AS1 in EBC1, A549 and MCF7 cells (p = 0.0005, p = 0.0109 and p = 0.0461 respectively), whereas L1MET_AS3 showed a lower effect, only evident in EBC1 cells (p = 0.0461) (Fig. 3). We observed a decrease of L1-MET expression also in MRC5 cells, however the basal level of L1-MET was too low to be informative. Moreover, normal lymphocytes did not express L1-MET (Fig. 3).
The cell viability assays were performed to investigate the effect of L1-MET silencing on cell growth. In line with the previous results, the L1MET_AS2 silencing caused a strong reduction in the viability of EBC1, A549 and MDA-MB231 cells (p < 0.0001) and a significant decrease in MCF7 cells (p = 0.028) (Fig. 4). The L1MET_AS1 silencing was significant in EBC1, A549 and MDA-MB231 cells (p < 0.0001, p = 0.0001 and p = 0.02, respectively), but not in MCF7 cells (p = 0.07). The use of L1MET_AS3 affected only the viability of EBC1 cells (p = 0.002) (Fig. 4). The viability of the control cells was not influenced by silencing L1-MET with any of the three LNA- GapmeRs indicating that they do not have a general toxic effect (Fig. 4).
To test whether the reduced cell viability could result from increased apoptosis, we evaluated the apoptotic index in the silenced cells by flow cytometry. In agreement with the previous results, we found that L1MET_AS2 silencing was associated with an increase in the apoptotic index in all cancer cells, with EBC1 and A549 cells displaying an average of 39.7% and 44.3% apoptotic cells, respectively (p = 0.0005 and p < 0.0001) (Fig. 5). In these two cell lines, L1MET_AS1 silencing showed similar effects (p < 0.0001 and p = 0.0007), whereas L1MET_AS3 did not affect apoptosis. MDA-MB231 and MCF7 cells underwent apoptosis when the L1-MET transcript was silenced using L1MET_AS2 (p = 0.0016 and p = 0.0014). As observed for the cell viability assay, MRC5 cells and human lymphocytes did not display alterations in apoptotic levels with any of the used LNA-GapmeRs, again excluding off targets effects (Fig. 5).
Role of the L1- MET transcript in the modulation of gene expression
The effect of L1-MET depletion on the modulation of gene expression was investigated by RNASeq analysis on RNAs extracted from A549, EBC1, MDA-MB231 and MCF7 cells silenced with L1MET_AS1, L1MET_AS2, and scrambled GapmeRs.
The hallmark of the cancer gene sets was investigated by GSEA (Supplementary Table 3). Overall, we observed that L1MET_AS1 and L1MET_AS2 treatments triggered a consistent positive apoptotic signal, as reflected by the upregulation of the P53 and TNFα pathways across all cell lines (Fig. 6A), generally associated with a negative signal in the cell cycle- related gene sets (mitotic spindle, G2M checkpoint). In detail, we found a uniform and specific downregulation of cell cycle genes (E2F, MYC and G2M) and oxidative phosphorylation pathways in all the analyzed cells upon depletion of L1-MET.
Differential Gene Expression (DGE) analysis evidenced that lung cancer cells (EBC1 and A549) had a considerably higher fraction of deregulated genes in respect to breast cancer cells (MDA-MB-231 and MCF-7) (Fig. 6B) (Supplementary Table 4). A significant decrease of the MET expression was found in EBC1 and A549 cell lines treated with both L1MET_AS1 and AS2, whereas a slighter MET transcript downregulation was evidenced in MDA-MB231 cells with L1MET_AS2. As expected, the faint detectable MET expression in MCF7 cells was slightly affected by both the treatments (Supplementary Fig. 2). Notably, among the most common cancer-associated genes, we detected a remarkable decrease of the EGFR gene expression in all the cancer cells, except MCF-7, upon transfection of L1MET_AS1. The same results were observed with L1MET_AS2 in EBC1 cells (Fig. 6C and Supplementary Fig. 2).
L1- MET effects on MET and EGFR protein expression
We characterized the potential downregulation of the MET and EGFR proteins in our cancer cell lines, together with their phosphorylation status and the effectors AKT and ERK. MRC5 fibroblasts were used as controls because of their very low level of MET mRNA expression.
The EBC1 cells, which are strictly MET-dependent for survival due to MET gene amplification and carriers of an activating variant of the EGFR, EGFR p.L858R, showed high expression of both MET and EGFR (Fig. 7). We detected reduced expression of both receptors and their corresponding phospho-proteins after the treatment with both L1MET_AS1 (MET p = 0.0018; EGFR p = 0.0137) and L1MET_AS2 (MET p = 0.0024; EGFR p = 0.0266). In line with these findings, we observed the loss of the AKT phosphorylation and the decrease of phospho-ERK with both L1MET_AS1 (pAKT p = 0.0347; pERK p = 0.0056) and L1MET_AS2 (pAKT p = 0.022; pERK p = 0.0011) (Fig. 7)
A549 and MDA-MB231 cells, which are MET and EGFR-independent for their survival, but express both the proteins at steady-state, did not show any constitutive phosphorylation of the two receptors and AKT. We detected a strong reduction of MET expression with L1MET_AS2 in both cell lines (A549 p = 0.0334; MDA-MB231 p = 0.0128), whereas L1MET_AS1 did not show any significant difference compared to the negative control. Conversely, EGFR displayed a lower expression in both the cell lines using either L1MET_AS1 (A549 p = 0.004; MDA-MB231 p = 0.0182) or L1MET_AS2 (A549 p = 0.0125; MDA-MB231 p = 0.0264). Differently from EBC1, A549 and MDA-MB-231 cells had no significant reduction of ERK and ERK phosphorylation (Fig. 7). In agreement with the fact that MCF7 breast cancer cells do not express MET and EGFR receptors, both LNA-GapmeRs had no effects on the expression and phosphorylation status of AKT and ERK. Similarly, no effects were found on the MRC5 fibroblasts which express very low levels of MET (Fig. 7)
{kind=link}