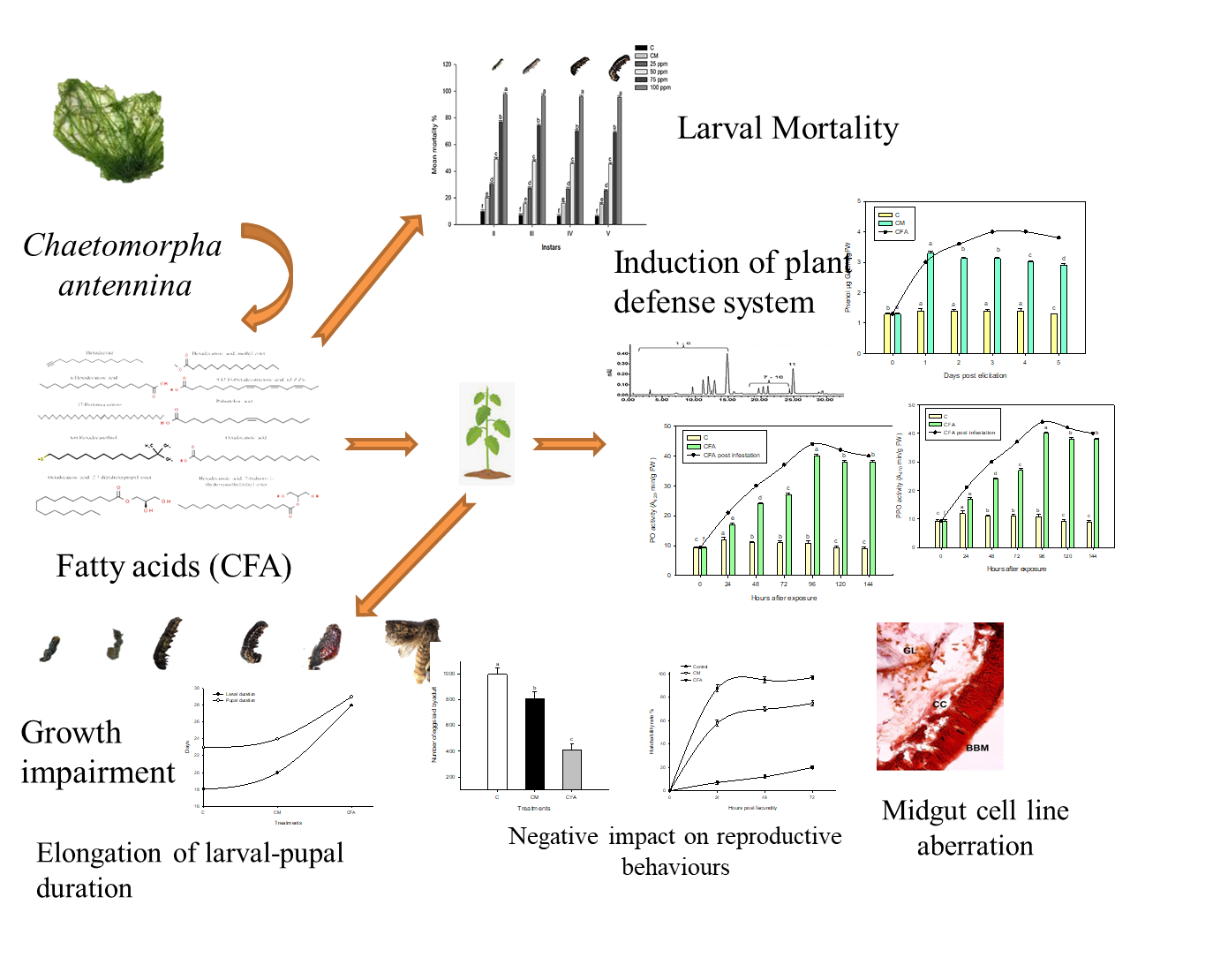
The predominance of the implication of green revolution practices that were once effective has attained a plateau. These methodologies relied on the use of high yielding disease resistant varieties, modified cultivation practices along with sumptuous application of chemicals for plant growth promotion as well as protection [36]. Considering all the negative impacts posed upon by the application of chemicals, there has emerged an immediate and alternative strategy to maintain the sustainability of agricultural production in a long run [37]. In this aspect, biofertilizers and biopesticides, sourced from natural materials are looked upon as an unconventional approach towards increasing the productivity. They are environmentally benign along with being safe, non-toxic and effective. Since these compounds are complex, the chances of development of resistance among the pest population are also significantly lower [3]. Seaweeds, which are superior to terrestrial plants in terms of bioactive chemicals, are used in culinary, medicinal, and industrial goods. Thus, the discovery of new botanical insecticides based on the different bioactive components of seaweed is critical. Since seaweeds are reported with higher weight/volume proportions of fatty acids, the possibility of testing their potentials as elicitors against the polyphagous pest, S. litura was analysed in this study.
Marine algae are regarded as excellent bases of assorted bioactive-compounds that can stimulate plant growing and also enhance resistance against environmental stressors. Further our experiment results do support the previous findings of Battacharyya et al. [38]. Fatty acids occupy a larger proportion of seaweed chemical composition. They are said to contribute to over 30% dry weight existing either as polyunsaturated or extractable fatty acids in green and brown algae [40]. Seaweeds have a high fatty acid (FA) variety, and many of them have potential bioactivity [40]. FAs are dynamic constituents of all biota, besides recognised for their imperative part in triggering the phytohormone interactions, apart from acting as key role players of various defense signalling pathways of the plant system [41].In the current study, C. antennina active fraction (CFA) displayed the presence of 19 fatty acids, with Hexadecanoic acid, Octadecatrienoic acid, linolenic acid, pentatriacontene and Octadecanoic acid in significant quantities. Hexa and octa decanoic acids are being reported with direct insecticidal activities. Fatty acids of Laminaria digitate, Undaria pinnatifida were reported with biocidal activities directly and offering protection by inducing innate defense system of plants in strawberry and lemon trees [42]. This was evident from the presence of salicylic acid and phenolic compounds in leaves of plants elicited with CFA.
The algal compounds induced the activities of PPO and PO, which are components of SA signalling pathways. Algal compounds have also increased the accretion of phenolic compounds in tomato leaves. Phenolic compound accumulation promoted by algal fraction application was significantly higher compared with cypmethrrin applications. Simultaneously, the treatments also offered effective protection against early blight disease and herbivory of S. litura. Algal treatments dramatically reduced the number of juvenile Meloidogyne incognita root-knot nematodes in soil while enhancing cumulative phenolic as well as antioxidant intensities [43]. The same results were obtained when commercial seaweed products Ecklonia maxima and Ascophyllum nodosum were used, which inhibited the reproductive as well as behaviour patterns of M. hapla and M. chitwoodi, that had infected tomato plants [44].
The elicitation test by CFA induced the production of SA which was evident from the HPLC chromatogram. Also SA was not observed in the chromatogram of control leaves. Hence the algal compounds were able to elicit plant’s systemic acquired resistance. Similar induction of SA by algal compounds was proved by Jaulneau et al. who reported the SA signalling pathway was induced by the application of algal polysaccharides, laminarin and carrageenans [45]. El Modafar et al. also proved ulvan compounds of Ulva lactuca stimulated SA-dependent systemic acquired resistance in tomato seedlings [18].
CFA also exhibited significant mortality rate against the larvae of S. litura (Instars II to V). Similar mortality properties of C. antennina phenols were demonstrated with a wide pesticidal activity against mosquitoes [46]. The seaweed extracts are more effective against mosquito larvae even at lower concentrations. This was demonstrated by Manilal et al., who showed that the extracts of various green and brown algae had a greater effect on the dipteran larvae, at lower LC50 values [40]. Sahayaraj et al. stated the efficacy of seaweed compound, tetradecanoic acid of Caulerpavera veravalensis were active against Dysdercus cingulatus nymph, a serious cotton pest [16]. It has been demonstrated that saturated FAs are abundant in seaweeds and are capable of killing agriculturally significant pests such as Sitophilus granarius [47].
The II instar larvae reared on elicited plants displayed extended larval duration due to reduction in feeding. Consequently, the biomass of these larvae was also very low. A similar larval period extension due to reduced feedant activity was observed in tobacco cut-worm larvae treated with Aristolochia tagala extracts [48], Momordica charantia secondary metabolites [49] and Citrullus colocynthis [50]. The extended larval periods drastically affected the pupae, reducing their biomass, size and duration. The extreme morphological damage to the pupa that metamorphosed from larvae with lower biomass and extended larval duration was also reported by treating Musca domestica larvae with parsley and citronella oil [51]. The pupal duration is the base for major nutrient and energy consumption required for the development of a healthy adult as well their fecundity and egg hatching rates [22]. The reduced pupal weight and duration also affected the further development of S. litura that extended the adult longevity, female fecundity and egg hatchability. A likely reduction in fecundity of females that emerged out of extended larval periods and lower pupal biomass was reported in S. litura larvae treated with leaf extracts of Momordica charantia [49]. Delayed metamorphosis moderates post-larval performances. An increase in larval duration increasingly posed a damaging effect on post-larval growth and survivability of Echinometra sp [52].
Plant metabolites are lethal to insect herbivores, by interfering with their food consumption and/or utilization. Food consumption is recognised as one of an organism's toxicological endpoints. [53]. The elicitation negatively affected the food consumption and consequently their nutrition. A reduction in the utilization of consumed food resulted in the lower growth rate of the larvae. This directly influenced the behaviour and physiology of the larvae post ingestion. An analogous effect on dietary utilization decline effected the development and physiology of rice leaf folder larvae treated with a biopesticidal combination [54] and Dysoxylum triterpenes [22].
Transphosphorylation processes hydrolyze phosphomonoesters by acid phosphatases in acidic settings and alkaline phosphatases in alkaline conditions. These actions are observed at a greater frequency in the insect midgut, the weakening of which will debar insect survival [37]. Current study reports the reduction in these enzymes, that was reported by previous research indicating the drop-down in such enzymes in insect pests treated with pesticides [55] and azadirachtin [22]. Reduced ACP-ALP enzyme activity correlates with low energy levels induced by metabolite transport disruption. In rice leaf folder larvae subjected to a biopesticide formulation including neem seed kernel and Bt toxin, a similar impact on ATPase enzyme activity was seen [37]. ATPase reduction was caused due to the cease of metabolism as a consequence of either food indigestibility or absence of food intake [56]. LDH enzyme plays a vital role in carbohydrate metabolism, are also indicators of chemical stress [22]. Reduced LDH levels in larvae developed on elicited plants is an indicative of lower carbohydrate metabolism as a result of reduced feedant activities. A likely reduction in LDH levels due to insect toxic allelo-chemicals in plant extracts were observed in neem limonoids treated rice leaf folder larvae [22]and S. litura larvae treated with and C. colocynthis extracts [50].
The digestive enzymes such as amylase, lipase and protease also displayed a downfall in larvae due to elicitor treatment. The overall decrease is due to the relative decline in the food consumption of the larvae. Similar decreases in digestive enzymes were observed in S. littoralis treated with extracts of Calotrophis procera [57]. The potential of insect larvae to detoxify the ingested toxic compounds depends on the efficiency of detoxifying enzymes such as GSTs, cytochrome P-450 and esterases. A reduction in overall activities of these enzymes in the larvae that fed on elicited plants signifies the inability of development of resistance by the insect pests against the defense compounds. A likely decline in the activities of detoxification enzymes were found in the insect larvae of Hyphantria cunea treated with Ginkgo biloba secondary metabolites [58]. Because larvae digest and absorb nutrients in the larval midgut area, histological study of the larval midgut was performed. The midgut histological study of treated larvae revealed a damaged brush boundary membrane, which might be owing to the active chemicals interfering with metabolite or ion transport. It's possible that this started a chain reaction of cellular processes that finally prevented the insect from eating, such as cell disintegration and the leakage of cellular components. This resulted in enlargement of the treated larvae's midgut area. Similar abnormalities in the midgut of S. litura treated with seaweed chemicals have also been documented [51].
Both the elicitor treatments increased foliar phenols, PO and PPO activities significantly higher compared with control. Increased production of phenolic compounds is considered as biomarkers of induced resistance [59]. Application of MeSA has stimulated the production of phenolic compounds [60]. Jasmonate application on tomato plants were found to stimulate the production of defensive proteins that negatively influenced herbivores [61].There exists a linear relationship between phenolic compound concentration and antimicrobial potentials [19]. Consequently, the allegation of phenol accumulation in plant defense has also been proved [62].
Plant mediated interactions amongst pathogens besides arthropod herbivores can ensure significant concerns for individual pests as well as their population dynamics. These types of interactions are mediated by SA and JA facilitated plant interactive pathways. While herbivore attack stimulates JA pathway, pathogen infection stimulates SA related defense signalling pathways. PO and PPO activities are components of early response in plants to pathogen infection and insect infestation [61].
Higher accumulation of foliar phenols and increased in the activities of enzymes PO and PPO by SA treatments are reported in Solanum melongena, Brassica juncea (var. Rlm619) that provided resistance against Ralstonia solanacearum [63]. Additionally, increased phenolics associated with amplified PAL activities conferred resistance against fungal phytopathogen, Fusarium oxysporum that was exposed with algal polysaccharides [18]. A similar resistance to fungal pathogens, Botrytis cinerea and Phytophthora infestans was observed by the treatment of tomato seedlings with algal products from Sargassum fusiforme [64]. The ability of algal compounds to offer better resistance compared with chemical pesticide, is attributed to the ability of these compounds to induce additional defense signalling pathways along with that of SA mediated defense responses.
A similar increase in PO and PPO activities were also observed with respect to elicitor treatments pre and post S. litura infestation. The levels were higher in infested elicited plants compared with that of un-infested. A likely increase in PRPs post infestation in elicited plants were also observed in tomato plants treated with commercial elicitors, commercial elicitors benzothiadiazole and methyl jasmonate post infestation with green peach aphid, Myzus persicae [65]. These elicitors also effectively reduced the aphid population and fecundity. The elicitor treatments induced plant PRPs in response to both pathogen and insect attacks. However, at 120h, the levels of PPO and PO were higher in response to herbivory. Nevertheless, Boughton et al. proved that the antagonistic reactions flanked by JA then SA reliant plant signalling conduits was able to deactivate infestation mechanisms of aphids [65]. Hence higher level of protection was conferred against insect infestation by the activation of both JA and SA pathways that resulted in the production of various secondary metabolites such as lignins, tannins, flavonoids, and enzymes such as PO, PPO, PAL and LOX. Stout et al. also stated the probable instigation of defense genes typically connected with pathogen attack [61].
{kind=link}