The diblock copolymer poly(tert-butyl acrylate)-block-poly(6-(3,6,7,10,11-pentakis(hexyloxy)-2-oxytriphenylene) hexyl methacrylate) (PtBA102-b-PHATMA22, Fig. 1a) was synthesized via sequential atom transfer radical polymerization (ATRP).18 The discotic HAT mesogen-containing monomer 6-(3,6,7,10,11-pentakis(hexyloxy)-2-oxytriphenylene)hexyl methacrylate (HATMA) and its homopolymer were synthesized according to literature (Scheme S1a, and 1b, Fig. S1-12, Table S1).19 The detailed molecular characteristics of the diblock copolymer were included in Scheme S1c, Fig. S13-17, and Table S2.
Although discotic LC polymers combine orientational molecular packing and fluidic nature and have been found attractive for thermal,20 optical,21 and electronic devices,22 its solution-state self-assembly behaviors have been rarely explored.23–26 The hexagonal columnar packing of HAT mesogens has been well understood. When they were attached as pendant groups for polyacrylates, the organization patterns were mostly retained,27, 28 but their LC properties were severely impaired, due to the limited mobility from the polyacrylate backbone.29 Very similar results were also obtained in the current study that the phase transition of the PHATMA block could not be detected via normal differential scanning calorimetry (DSC) characterization (Fig. S18). The LC ordering effect was so weak that the orderedness could not be restored within a reasonable time range during the cooling process. To observe the LC to amorphous phase transition, we had to anneal the sample at 35 oC for 24 h before the DSC characterization and record the results from the first heating scan (Fig. S19). However, this weak LC ordering effect makes it possible to set up a delicate balance between different interactions during the micellization process, triggering the continuous morphological transitions over time.
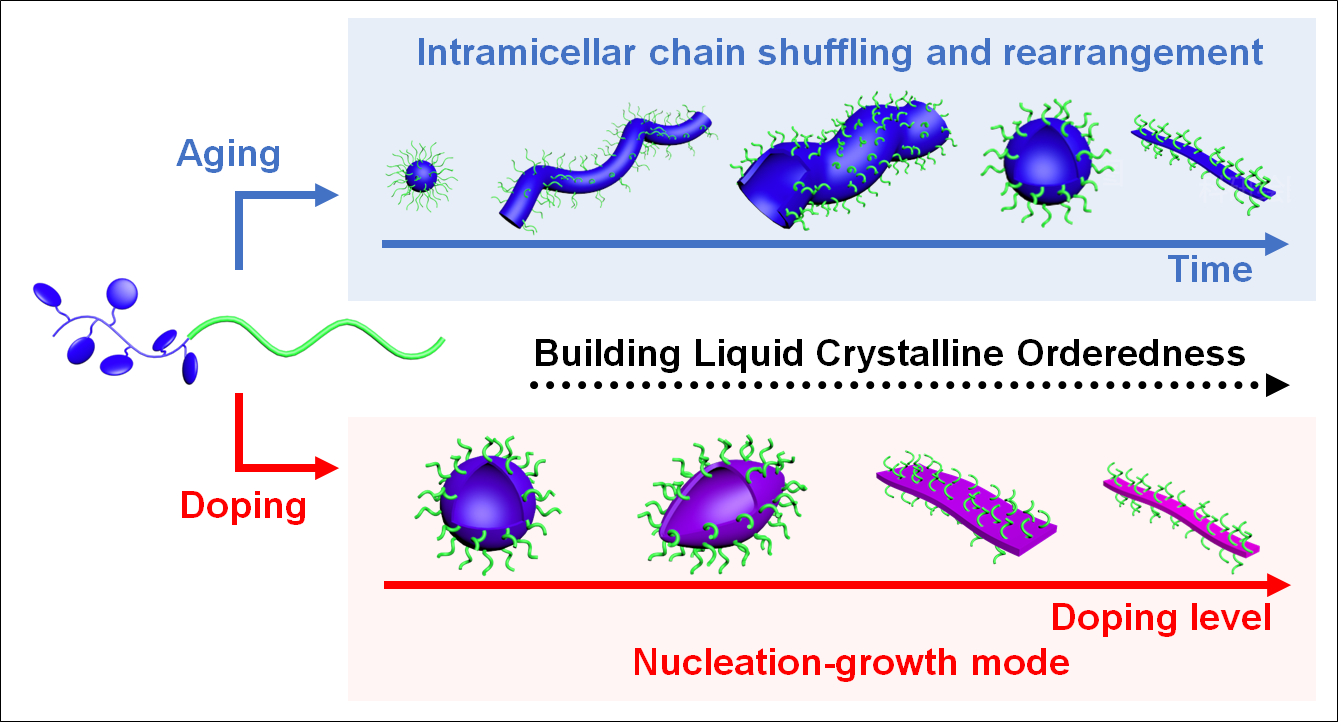
For the self-assembly experiment, the diblock copolymer was directly dispersed in 2-propanol (2-PrOH, concentration = 0.1 mg/mL) at 80 oC for 1 h and cooled down naturally to room temperature (r.t., 21 oC) within 3 h. Intricate multiple micellar morphological transitions were subsequently observed (Fig. 1b upper) via transmission electron microscope (TEM). Aliquots were taken at different stages and dried on TEM grids within seconds before observation (Fig. 1c-1j). Initially, at 80 oC, all the polymer was dispersed (Fig. S20). When the solution was cooled down to 75 oC, spherical aggregates appeared (Fig. 1c). At 60 oC, these spherical aggregates fused into thick worm-like aggregates with a diameter of around 100 nm (Fig. 1d). After the solution reached r.t., these worm-like aggregates were further swollen into tubular structures with dramatically increased diameter (~ 500 nm, Fig. 1e). Interestingly, after 2 hours at r.t., these tubular structures started to divide into vesicles (Fig. 1f), of which the process would finish within another 10 h (Fig. 1g). Moreover, these vesicles started to spin out thin fibrils after 120 h of storage (Fig. 1h). These jellyfish-like structures transformed into thin fibrils exclusively in 120 days (Fig. 1i), for which a diameter of 25.6 ± 4.6 nm, and a height of 2.0 ± 0.1 nm were measured from their TEM and atomic force microscopic (AFM) images (Fig. 1j), respectively.
It is not too surprising for BCPs to undergo one or two morphological transitions spontaneously during self-assembly,30, 31 but it has been never reported that so many distinct morphologies could be shown in a single self-assembly process. A plausible driving force would be the subtle LC ordering effect from the HAT mesogens. To better understand its role during the morphological transition, the fluorescence (FL) emission of HAT moieties was first monitored to look for a hint from their molecular packings (Fig. 2a). After the solution reached r.t., the peak intensity at 390 nm (excitation wavelength = 270 nm) gradually decreased over time until 120 h, due to the enhanced π-π stacking from the slowly organizing HAT mesogens.32 Afterwards, with the gradual formation of fibrils, the peak shifted toward a longer wavelength, due to the even stronger π-π stacking,33 indicating the further boosted orderedness of the discotic mesogens.
The wide-angle X-ray scattering (WAXS) spectra of the pure vesicles and fibrils were also obtained to explore the molecular packing of the mesogens. Both spectra showed hexagonal columnar packings of the HAT mesogens (Fig. 2b). However, the (110) peaks were not as obvious as they were from homopolymers (Fig. S12), due to the strong disturbance from the amorphous PtBA block.34 Closer examinations of the spectra revealed a more obvious peak for the thin fibril sample at q = 1.85 Å−1, corresponding to the (001) plane of the columnar phase, suggesting a higher LC orderedness for the thin fibrils. Moreover, the peaks at q = 0.17 Å−1 and 0.34 Å−1, from the lamella packing of the columnar mesogens, also appeared to be more obvious for the fibril sample. Meanwhile, with the morphological transformation from vesicles to fibrils, the lengths of the a-axis decreased from 21.33 to 20.9 Å, and the d001 also decreased from 3.53 to 3.40 Å (Fig. S21, and 22). All these results suggested a higher LC orderedness for these fibrils.
The grazing incident wide-angle X-ray scattering (GI-WAXS) results showed that the HAT disks were stacked perpendicular to the vesicular membrane (Fig. S23). Meanwhile, the orientational LC packing of discotic mesogens within the core of these thin fibrils was verified via infrared-AFM (AFM-IR).35 Topographic image (Fig. 2c) of the thin fibrils were first obtained, and AFM-IR spectra (Fig. 2d) were acquired with the IR beam both parallel (Iparallel) and perpendicular (Iperpendicular) to the fibril at the marked black spot in Fig. 2c (Supplementary Note 1 and Figs. S24, and 25).36 At the wavenumber of 1435 cm− 1, which corresponds to the in-plane bending vibration of C = C bonds on the HAT disks, the dichroic ratio (DR = Iparallel/Iperpendicular) values reached 3.3 (Fig. 2d), demonstrating that the discotic mesogens were preferably packed in columns perpendicular to the length of fibrils.35 Therefore, these fibrillar micelles appear more likely very thin belts.
Consequently, a detailed route for the morphological transition on the molecular level could be proposed. Initially, spherical aggregates were formed to lower the interfacial energy, due to the reduced solubility of the PHATMA block. The HAT moieties were amorphous in the micellar core. Subsequently, the discotic HAT disks started to slowly pack into ordered structures. At this stage, the orderedness of the disks inclined over time, and the density of the micellar core should increase and its volume decrease spontaneously. For the micellization process of coil-coil BCPs,37 this change should prefer the formation of spherical micelles over worm-like micelles. However, our results showed exactly the opposite trend. This counterintuitive observation clearly demonstrated that the subtle LC ordering effect from HAT moieties played a crucial role in the micellization process. During the next stage, the hexagonal columnar phase inside the vesicular membrane continued to pack even more closely. This would increase the crowdedness of the coronal chains, causing the further transformation from vesicles to fibrils (Fig. 2e).31
It is noteworthy that if the solution was quenched to r.t. within 1 min, the HAT mesogens were not able to organize into a well-defined columnar LC phase (Fig. S26), resulting in random aggregates (Fig. S27a). The poor flexibility of the PHATMA chains hindered the LC ordering process, and these random aggregates would not evolve into well-defined structures even after 6 months of storage (Fig. S27b), demonstrating the vital role of the LC ordering effect in the morphological transition process.
Another especially interesting point for the LC ordering process is that the orderedness not only can be gradually built over time via intrinsic LC ordering effect, but also strongly enhanced instantly by mixing them with appropriate dopants.38, 39 Particularly, it has been reported that HAT mesogen can be doped with TNF (Scheme S2, Fig. S28) to form electron donor-acceptor complex (EDA), and significantly enhance the orderedness and thermal stability of the LC phase.40 In the current case, when TNF was added to the tetrahydrofuran (THF) solution of PHATMA, the colorless solution immediately turned black, due to the formation of EDA (Fig. S29). The formation of EDA was confirmed with various spectrometric analyses as well (Fig. S30-32). Therefore, for the following assembly experiment, the dopant was added to the THF solution of the diblock copolymer initially to allow them to fully complex with HAT mesogens, and the sample was dried under nitrogen flow before thermal dispersion into 2-PrOH.
To investigate the influence of doping level on the assembly morphology, we first designed a complicated experimental procedure to show that the dopants were complexed with the diblock copolymer quantitatively (Supplementary Note 2 and Fig. S33). Therefore, the doping level (r = mole of TNF/mole of HAT) could be tuned quantitatively. By increasing r from 0.01 to 1, not only did the isotropic to LC phase transition temperature increase (Fig. S34, and 35), but also the d-spacing decreased accordingly (Fig. S36, and 37), suggesting the enhancement LC orderedness with the increase of r.
The micellar morphologies were substantially influenced by doping (Fig. 3). With very small r-values (0.01 to 0.05, Fig. S38), the spherical vesicles (Fig. S39) would not completely transform into fibrils, and this jellyfish-like morphology was retained even after one-year storage at r.t. (Fig. S40). Interestingly, when r-values increased over 0.05, elongated vesicles were obtained (Fig. 3a and Fig. S41), which were formed via the deformation of spherical vesicles during cooling (Fig. S42-44), and their aspect ratio increased with r-values to above 2 (Fig. S45). Their morphological transition was also halted during the spinning of fibrils (Fig. S46). With the increasing doping level, the rigidity of the LC PHATMA domain also increased, these vesicles had to elongate and release the bending penalty by forming topological defects at the vertices of the vesicles.41, 42
When the r-values reached 0.3, the membrane became too rigid that it could not be confined into closed vesicular structures, but instead flat bilayer structures were preferred to accommodate the high bending penalty of the mesogenic phase. As shown in Fig. 3b, and Fig. S47, broken vesicles were observed, from the edge of which some belt-like structures stretched out. With further increasing of r values, the assemblies would transform toward belt-like structures, which were exclusively observed when r reached 0.5 (Fig. 3c and Fig. S48). More interestingly, most of these belts appeared to be bent instead of straight, which has been rarely observed.43 Additionally, some rod-like structures could be visualized within the belt-like structures, as indicated by the white arrows in Fig. 3c. With further increase of r, the belt-like assemblies transformed toward fibrils (Fig. 3d and Fig. S49).
Surprisingly, when r = 1, exclusive fibrillar structures were obtained immediately after the solution was cooled to r.t. (Fig. 4). These fibrils appeared to be similar to those obtained from the undoped BCP, except that these fibrils were significantly thicker than the undoped ones, with a diameter of 45.1 ± 7.0 nm (Fig. 4a) from TEM images and a height of 4.0 ± 0.3 nm (Fig. 4b) from AFM images. Closer examinations revealed that the d-spacing of the doped fibrils (3.34 Å, Fig. S50) was also smaller than that of the undoped ones (3.40 Å), suggesting a higher LC orderedness (Fig. 4e, Fig. S51). With doping, the TNF molecules were inserted between HAT discotic mesogens to form an alternatively-layered structure, leading to thicker fibrils. Additionally, the higher LC orderedness inside these doped fibrils also led to an obviously higher rigidity, as indicated by their larger persistence length (14048 ± 332 nm) than that of the undoped fibrils (4405 ± 149 nm) (Fig. S52, Supplementary Note 3).44
The enhancement in LC orderedness within the micellar core induced by TNF doping could be confirmed with the WAXS results (Fig. S53), which agreed with those from TNF-doped PHATMA homopolymers (Fig. S36, and 37). This unambiguously confirmed the addition of a dopant could markedly enhance the LC orderedness, which plausibly drove the formation of these unusual micellar morphologies (Fig. 1b lower).
It has been well established that the orientational alignment of discotic mesogens would be beneficial for electronic devices,45 and the formation of EDA could further enhance the conductivity.46 As a proof of concept, the conductivity of the doped and undoped fibrils in the z-axis was measured with conductive AFM (c-AFM).47 As shown in Fig. 4c, the doped fibrils showed obviously enhanced conductivity compared to the undoped fibrils, confirming our hypothesis.
Moreover, it was found strangely that initially at high temperatures, instead of forming multiple morphological transitions, short rods were observed (Fig. S54a). With slightly lower solution temperatures, the rods started to grow rapidly in length, resulting in long fibrils eventually (Fig. S54b-54h). The lengths of these fibrils were reasonably uniform and could be measured before they became too long after the solution temperature dropped below 50 oC (Fig. S54). By calculating their number-average lengths (Ln), weight-average lengths (Lw), length polydispersity (PDI = Lw/Ln), and plotting the Ln and PDI with temperature, a typical exponential growth curve was observed (Fig. 4d, Table S3). Meanwhile, their PDI remained quite low during the growth process. This morphological evolution process closely resembled the in-situ nucleation-growth process,48–50 completely different from the undoped case.
Subsequently, based on such nucleation-growth assembly mechanism, these doped fibrils were subjected to “self-seeding” procedures to produce highly uniform fibrils. These pristine fully doped fibrils were first fragmented via ultrasonication at 0 oC to yield short rods (seeds, Fig. S55). The Ln was determined to be 121 nm from their TEM images. Seed solutions (total concentration = 0.005 mg/mL) were thermally annealed for 1 h at desired temperatures from 30 oC to 72 oC, and the annealed solutions then were cooled naturally to r.t. The rods with lower LC orderedness dissolved upon heating, while those with higher orderedness survived and functioned as seeds for the dissolved chains to grow epitaxially during the cooling process.49 With an inclining annealing temperature from 30 oC to 72 oC, the fibrils with narrow length distributions (PDI ≤ 1.04) were obtained and their lengths increased accordingly (Fig. S56; Table S4). Typical TEM images for the samples annealed at 65 oC (Ln = 270 nm, PDI = 1.03) and 72 oC (Ln = 611 nm, PDI = 1.02) were included in Fig. 5a, and b. The length information for samples at different annealing temperatures was summarized in Fig. 5c. The fraction of surviving seeds at each annealing temperature was calculated as well (Supplementary Note 4). As shown in Fig. 5c, in the range of 60–72 oC, the fraction of surviving seeds decreased exponentially with increasing temperature, a key characteristic of a self-seeding process (Fig. 5e, Supplementary Note 5, Fig. S57).51 Interestingly, after being aged for a long period, a large portion of these fibrils underwent end-to-end coupling to form segmented supramolecular micelles (Fig. 5d, and 5e), similar to our findings in other LC BCP systems.52 In sharp contrast, for the undoped fibrils, heating only led to their dissolution and restart of the morphological transition (Fig. S58).
Consequently, a plausible explanation for the formation of bent belt-like structures could also be proposed. Similar to the case of thick fibrils, short rods were observed initially at 80 oC (Fig. S59a), due to the existence of highly doped diblock copolymers and the formation of a highly ordered mesogenic phase. These short rods functioned as seeds to trigger the growth of the less-doped BCP chains during the cooling process (Fig. S59b). The unsaturated doping caused the nonuniform distribution of rigidity and stress inside the micellar core, eventually leading to the formation of bent belts. Similarly, self-seeding procedures could be applied to belt-like structures to obtain uniform belt-like structures (Fig. S60), strongly demonstrating the nucleation-growth mode again.
However, it was noteworthy that if TNF was added to the micellar solution, the morphological transitions would not be influenced (Fig. S61). Even with the excessive TNF (r = 10), the vesicular micelles still slowly evolved into jellyfish-like structures, and eventually thin fibrils within the same range of time as those without doping. This demonstrated that TNF molecules were not able to enter the mesogenic micellar core if they were not complexed with HAT prior to the assembly process. Therefore, such morphological transition should occur via the intramicellar chain shuffling and rearrangement, without involving individual BCP chains and dissociation/reassociation of the assemblies.53 On the contrary, with doping, the formation of EDA complexes completely changed the assembly mechanism to nucleation-growth mode. The morphological transition was no longer a result of the balance between different energy items but was dictated by the LC ordering effect. Such doping-induced alteration of the assembly mechanism was unprecedented.
Lastly, some comprehensive understandings can be concluded on the solution self-assembly behaviors of this discotic mesogen-containing diblock copolymer. As shown in Fig. 6, the energetic landscape and pathway of this assembly system are highly convoluted. The LC orderedness can be achieved in two different approaches. Due to the subtle intrinsic LC ordering effect from this PHATMA block, the LC phase can be slowly built over time. Consequently, the micellar morphology results from the delicate balance between different energetic items, such as the configurational entropy change of the core block after compression in the micellar core, the interface energy reduction via micellization, and the energy lost by stretching of the corona block,31 leading to slow morphological transitions to reveal multiple metastable micellar morphologies. Meanwhile, the LC ordering effect can be significantly enhanced upon doping to strongly influence or even dominate the morphological transition. With low doping levels, dopants will cause local orderedness variations and interfere with the morphological transitions, yielding exotic morphologies. With high doping levels, due to the uneven distribution of dopants, the highly-doped diblock copolymer can firstly aggregate to form mesogenic domains, which subsequently function as seeds to trigger the growth of the less-doped polymer chains. Finally, if the mesogens are fully doped, thick fibrils will be formed via a typical in-situ nucleation-growth mode, driven by the enhanced LC ordering effect. Such a doping-induced route to enhance the organization ability of assembly motifs presents a novel, versatile, and efficient strategy to achieve precisely controllable assembly from systems with weak ordering effects. An external component is added to enhance the organization ability of the assembly motifs, of which the underlying rationale is quite similar to the controllable assembly of supramolecular motifs based on guest/host pairs.48, 54 We envisage that it can be widely applied to many other assembly systems and some interesting features such as chiral assemblies can be readily realized via chiral doping.55
{kind=link}