Species of B. subtilis have the potential to produce two dozen antimicrobial substances, and 5–8% of the B. subtilis genome contributes to the production of antimicrobial substances [33]. Some inhibit the growth of pathogens and the germination of spores. The lipopeptide mixture of B. subtilis C232 inhibits the formation of Verticillium dahliae microsclerotia [34], and the volatile compounds secreted by B. subtilis JA inhibit the conidial formation and mycelial growth of Glomus etunicatum[35].
However, certain bioactive compounds are synthesized only under special conditions or as the result of external stimulation; therefore, it is difficult to obtain all the antimicrobial compounds produced by Bacillus using traditional cultivation and extraction methods, and this limited the comprehensive understanding of the mechanisms of biological control and biocontrol bacteria [22]. Genome mining allows the prediction of metabolites based on genome sequences and is widely used in obtaining new antibiotics [26]. It was used to identify a new NRPS pathway product, coelichelin, in Streptomyces coelicolor [36]. Pseudomycoicidin in Bacillus pseudomycoides DSM 12442 was discovered through the heterologous expression of its BGC in Escherichia coli[37]. Traditional cultivation and extraction methods were used to identify lipopeptide, fengycin, and surfactin from B. subtilis NCD-2, and fengycin showed strong antifungal abilities against V. dahliae and B. cinerea. However, the fengycin-deficient mutant of strain NCD-2 still has a certain antifungal ability, but it is less than that of wild-type strain NCD-2. Therefore, other antifungal active compounds, besides fengycin, may be produced by strain NCD-2 [2, 28, 29]. In this study, genome mining was conducted to analyze the potential antimicrobial compounds of the NCD-2 strain, and some of them were identified using MS. In total, nine kinds of secondary metabolite gene clusters related to surfactin, bacillaene, fengycin, bacillibactin, subtilosin, bacilysin, two terpenes, and one unknown product were identified from the genome of strain NCD-2. The surfactin [38], bacillaene [39], fengycin [40], bacilliactin [41], subtilosin [42], and bacilysin [43] showed antimicrobial abilities and played different roles in suppressing plant diseases. Only fengycin and surfactin were identified from the lipopeptide extract of NCD-2 despite the presence of other gene clusters in the genome. These other antimicrobial compounds may not have been detected because the acid precipitation extraction method was not suitable. Some bioactive compounds, such as bacillaene, bacillibactin, subtilosin, and bacilysin, are not lipopeptides. Therefore, these substances were not extracted using hydrochloric acid precipitation [44–47].
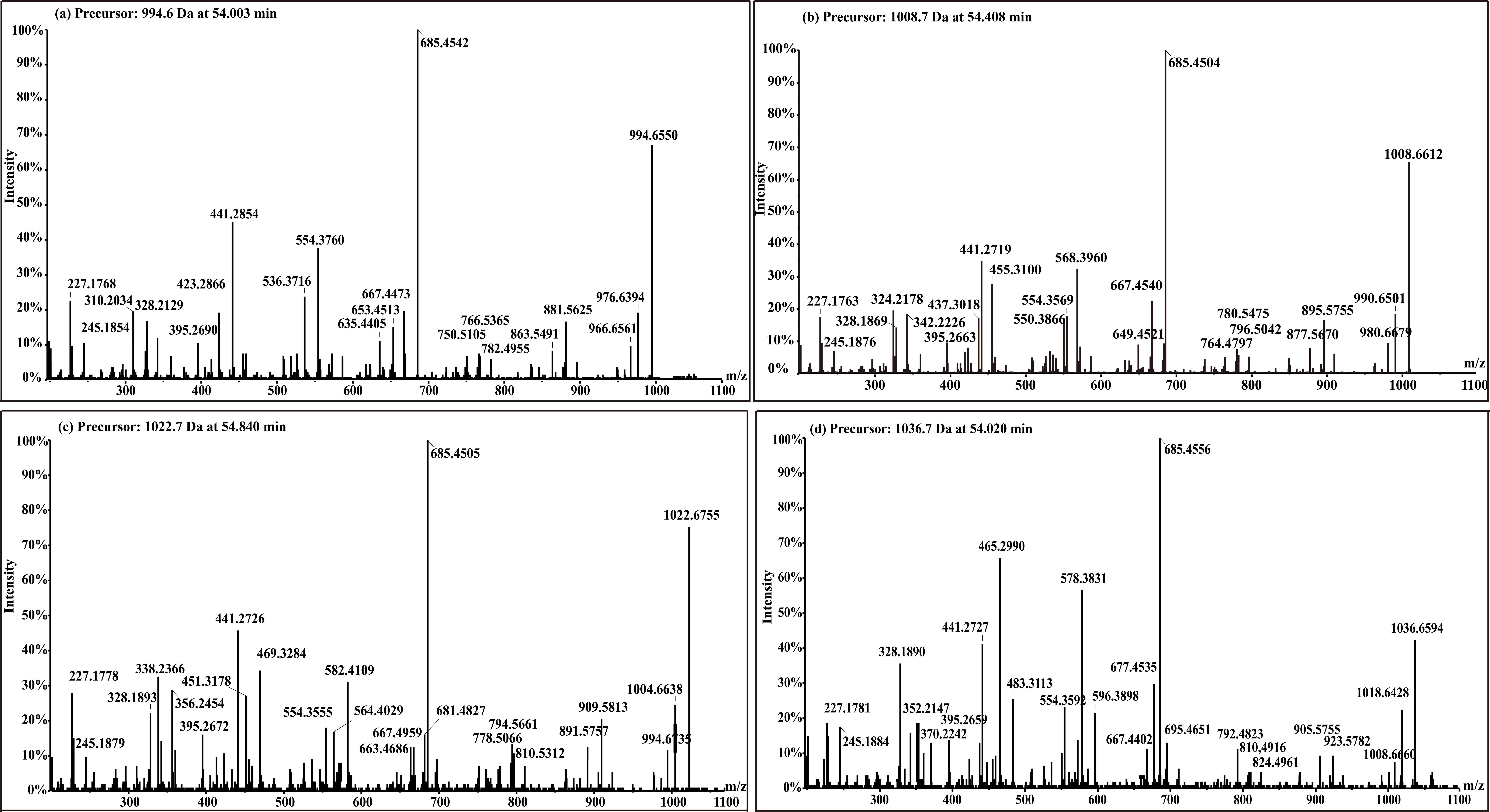
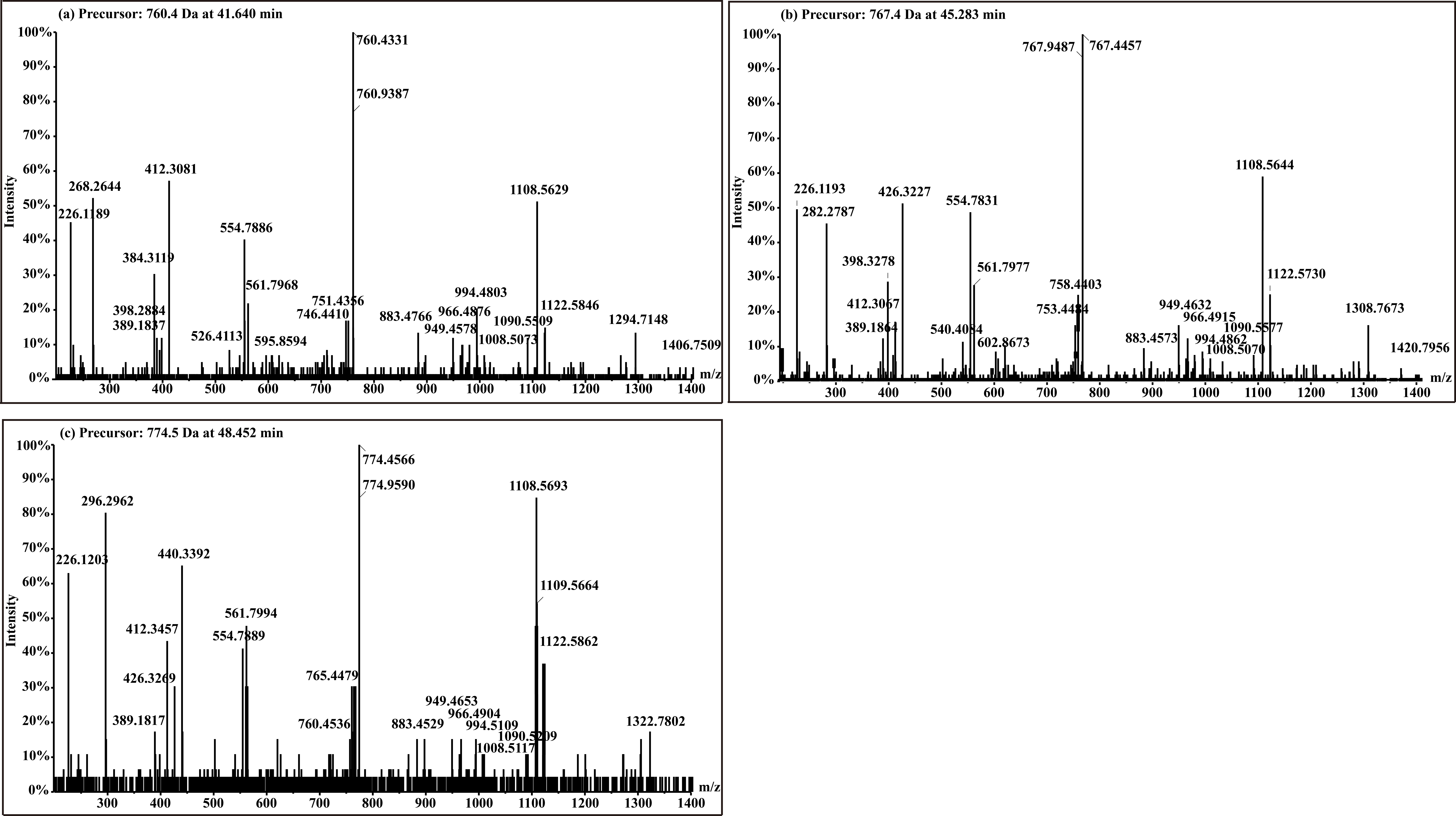
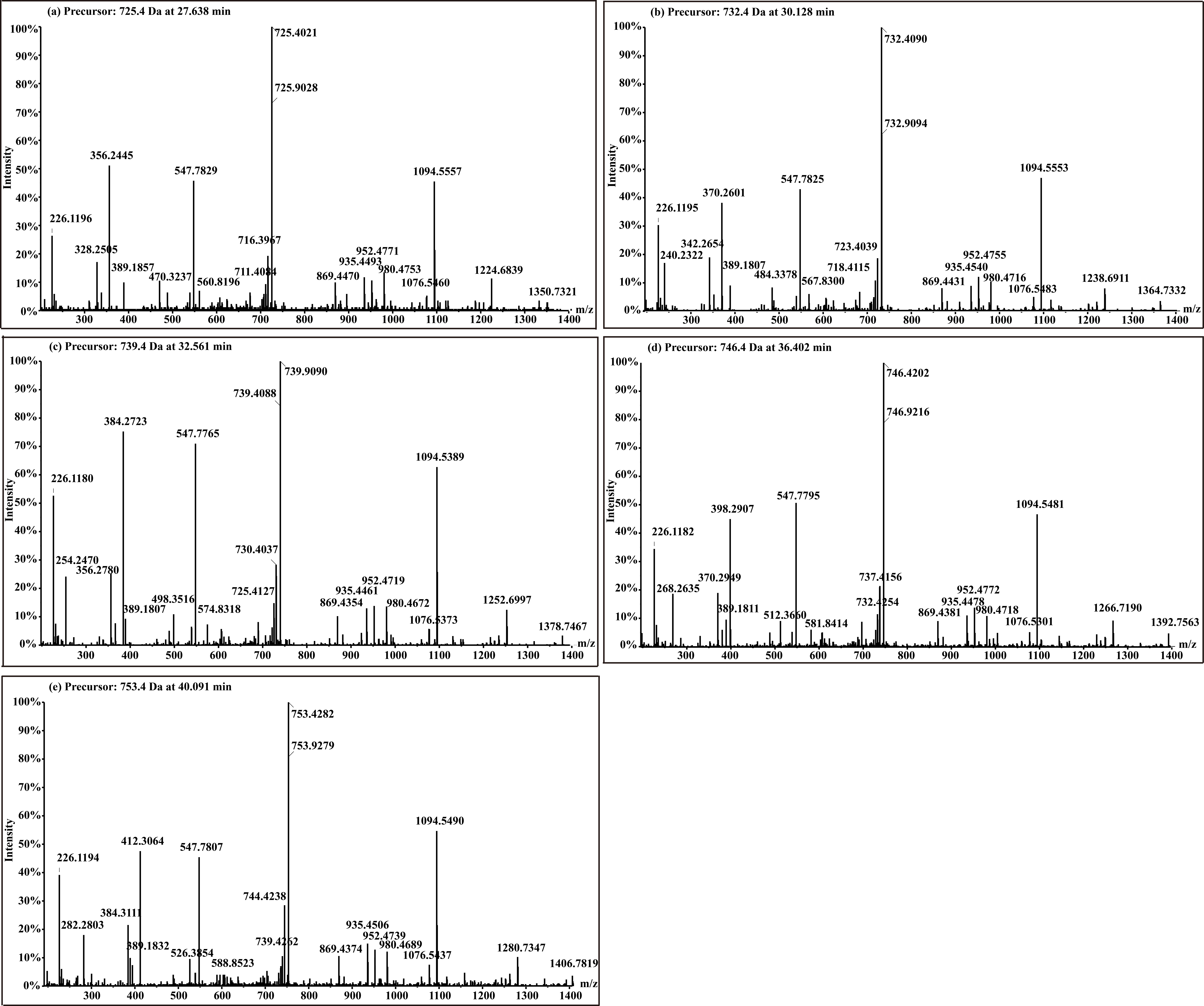
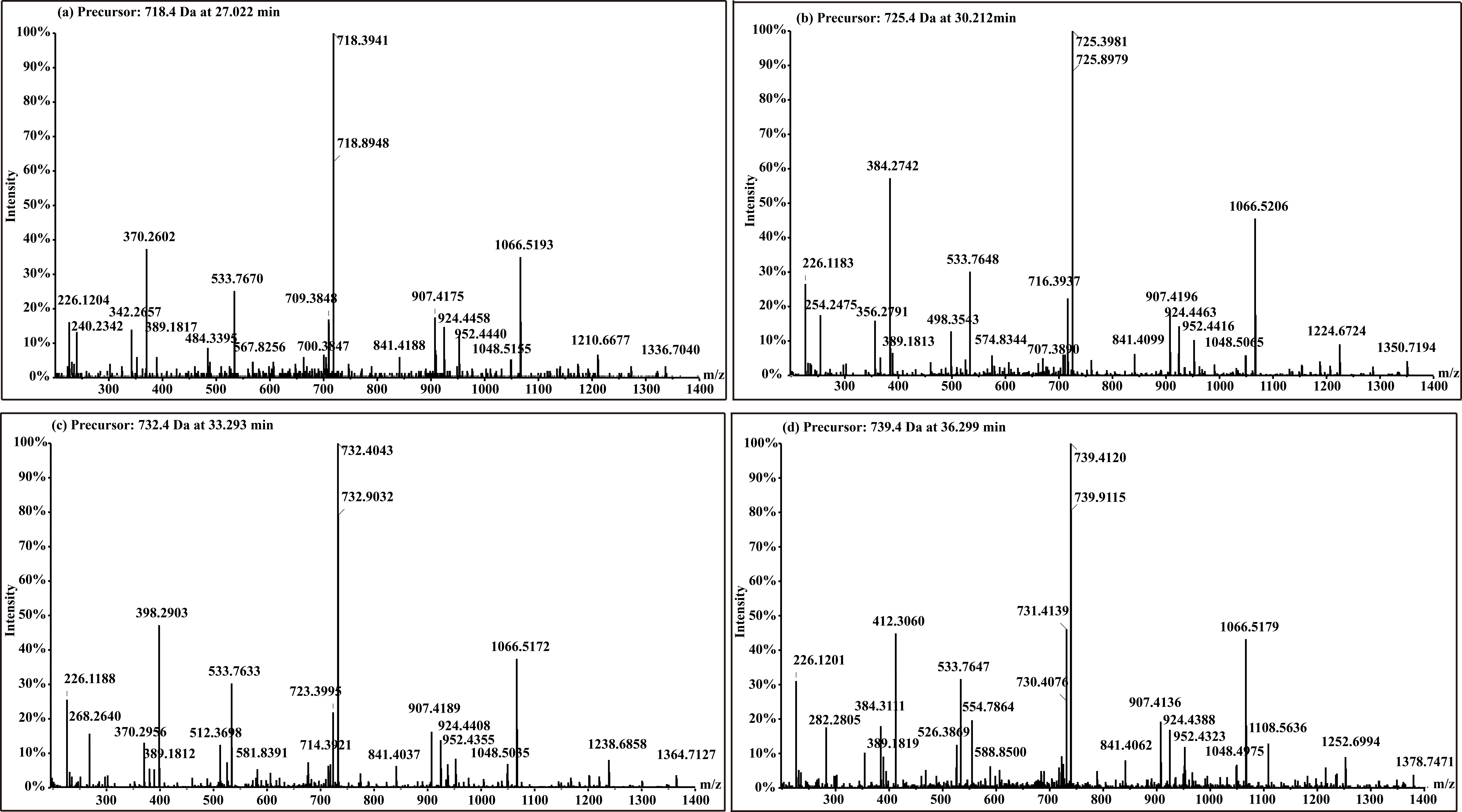
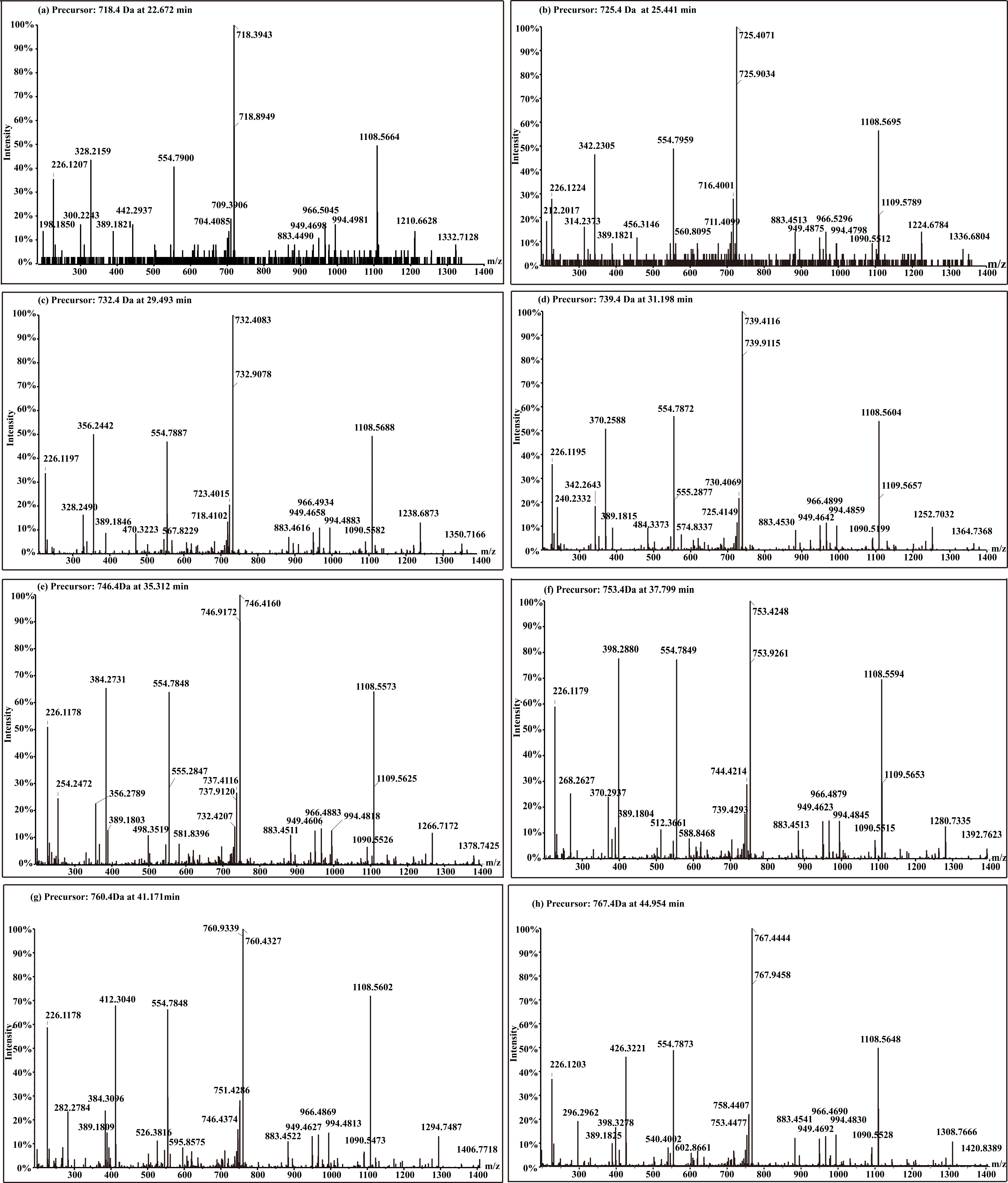
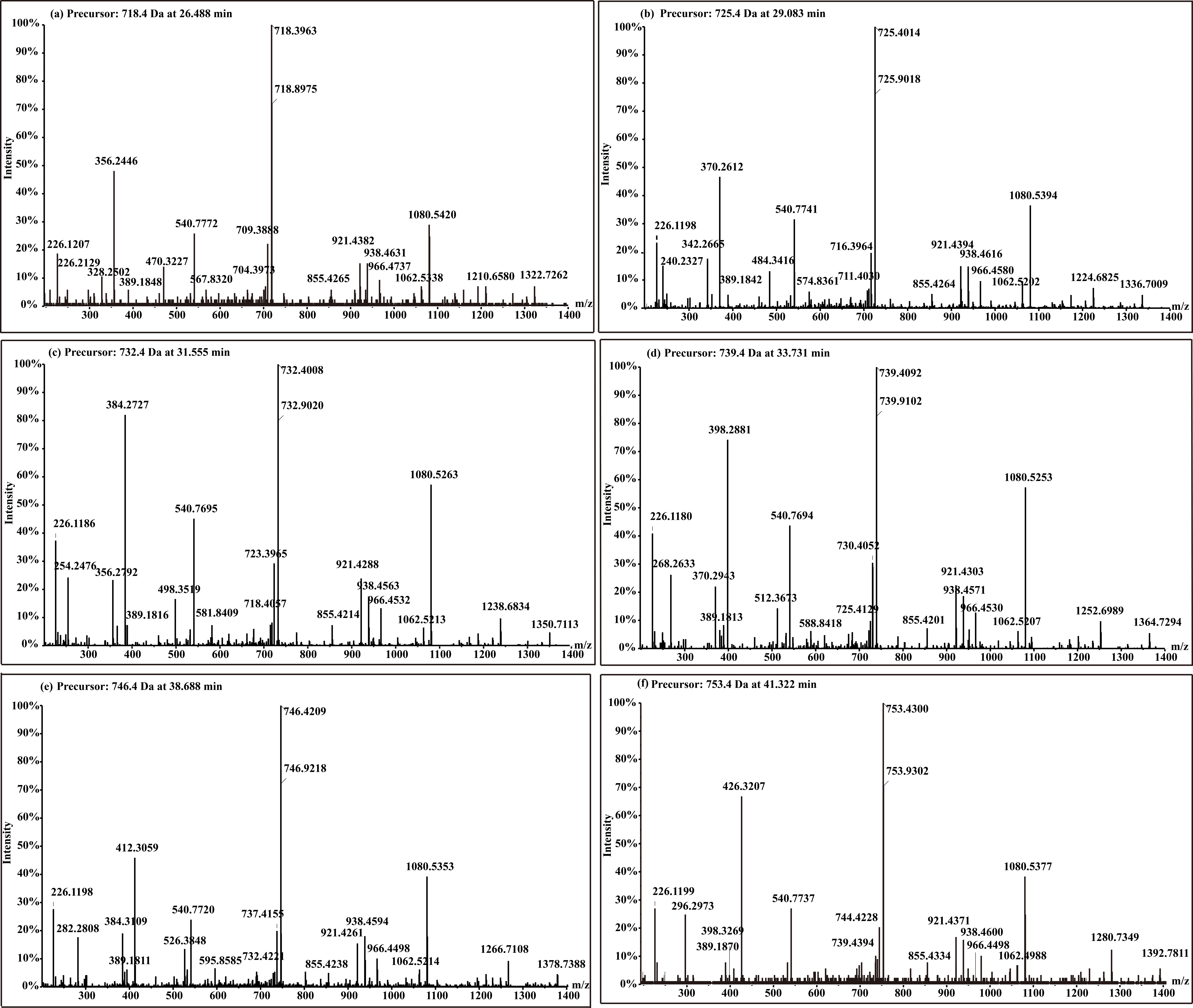
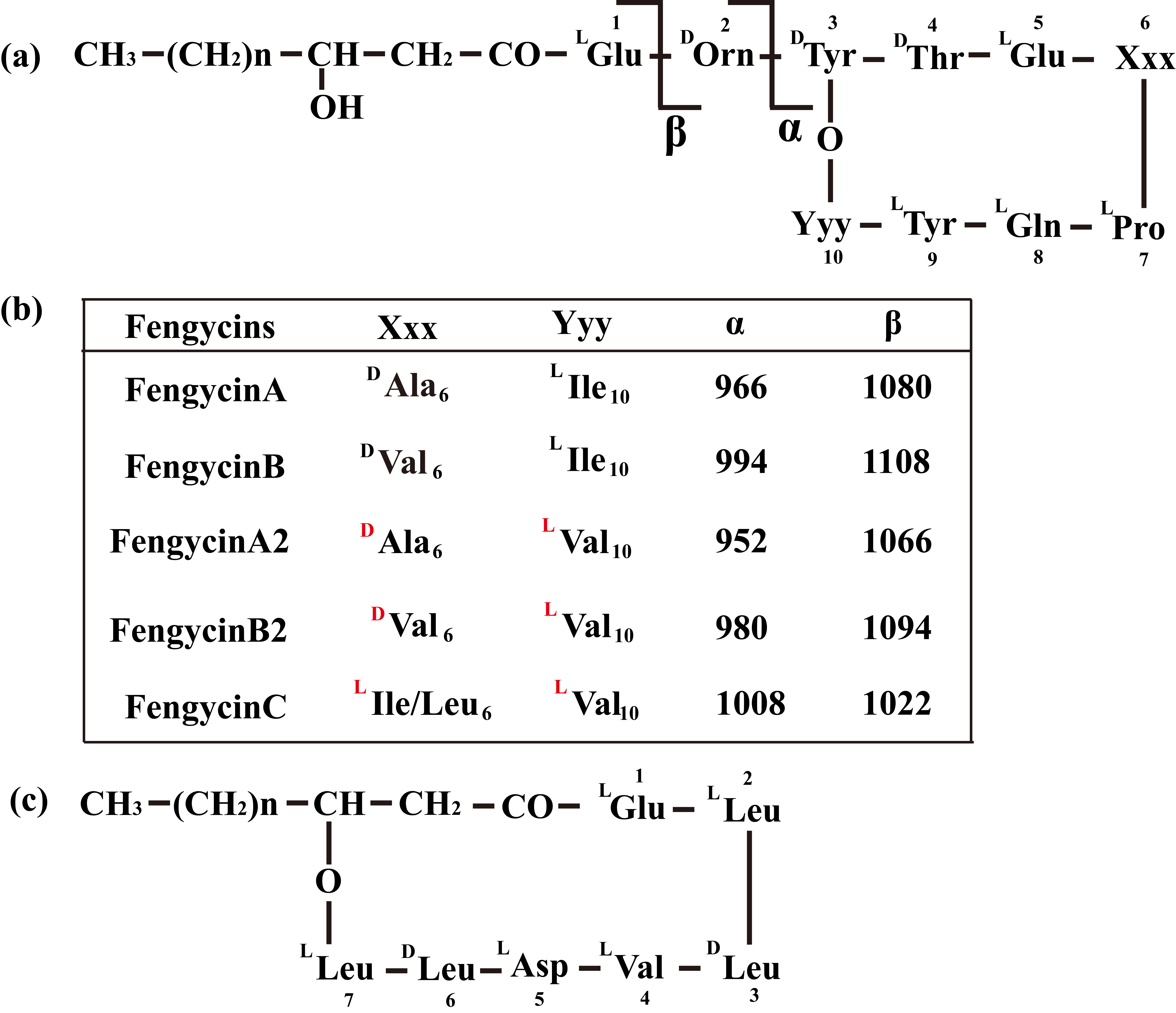
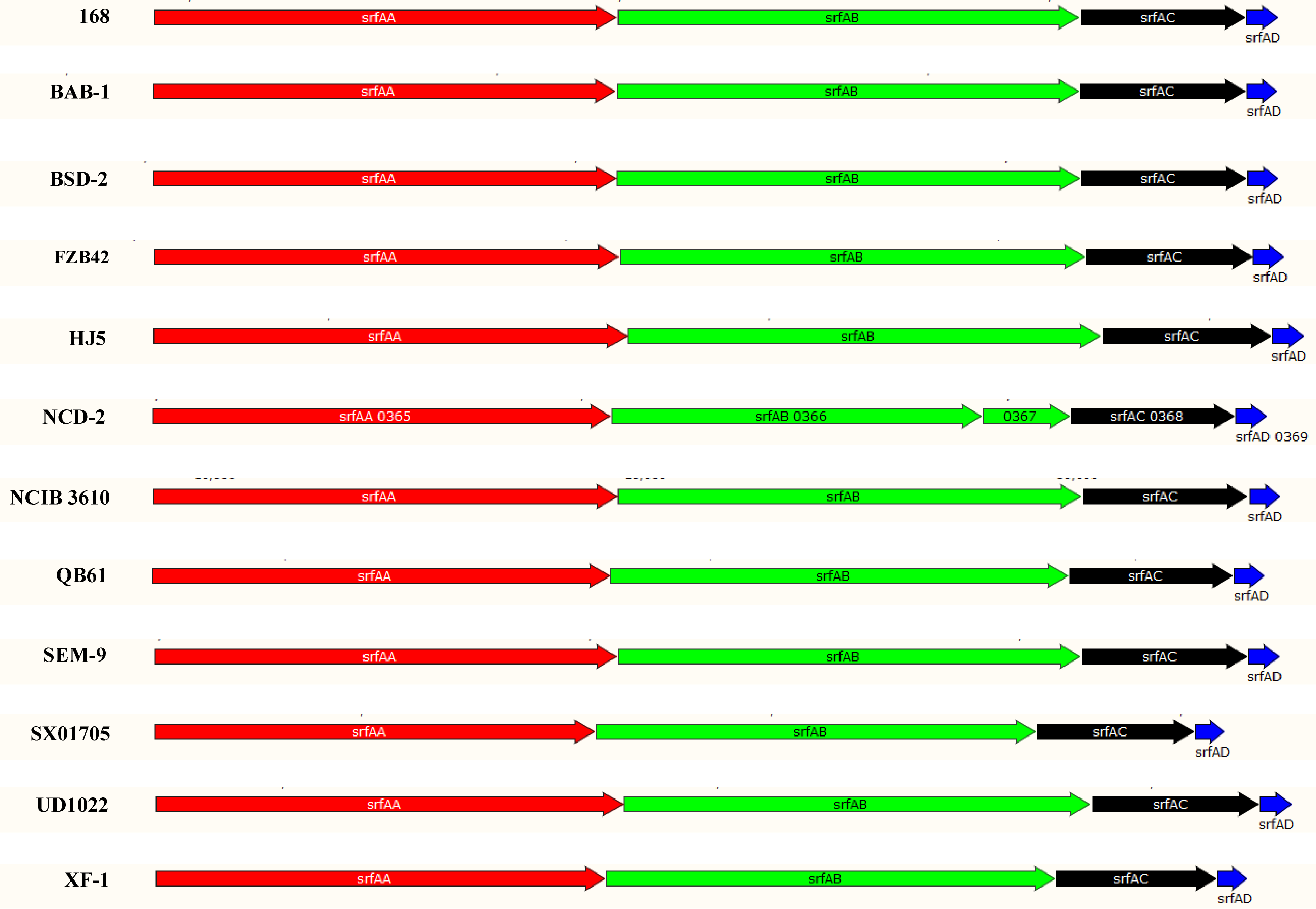
Fengycin comprises a peptide ring circled by 10 amino acids with a fatty acid chain tail. The mechanism of fengycin synthesis has been well studied in B. velezensis strain FZB42 [48], and the fengycin synthetic gene cluster in the strain consists of five genes (38 kb) that encode the synthetases FenCDEAB, of which FenC recognizes and carries glutamate and ornithine, FenD recognizes and carries tyrosine and threonine, FenE recognizes and carries glutamate and valine, FenA recognizes and carries proline, glutamine, and tyrosine, and FenB recognizes and carries isoleucine. FenCDEAB recognizes 10 amino acids and carries them to the β-OH FA chain to form fengycin [49–51]. However, NCD-2 only had fenEAB, lacking fenC and fenD, compared with the typical cluster structure of fenCDEAB in the FZB42 strain and 10 other Bacillus strains (Fig. 4b) and (Additional File 1,Fig. S1). To identify the enzymes FenC and FenD in the NCD-2 genome, their amino acid sequences from FZB42 were selected to screen for homologs by scanning the local NCD-2 proteome using BioEdit. The Gms1961 protein in the NCD-2 strain had the greatest similarity to FenC at an amino acid sequence level (Additional File 1,Table S2). The Gms1961 protein contained 2,550 amino acids, and the molecular weight was 287.50 kDa. The substrate bound by the adenylation domain of the Gms1961 protein was predicted (Additional File 1, Table S4). The adenylation A9 domain bound valine and N5-hydroxyornithine, with the latter being a transitional form of ornithine combined with the adenylation domain [52]. The UHPLC-QTOF MS/MS of the fengycins revealed that all the structures possessed the amino acid ornithine at position 2 (Fig. 5a–e), indicating that there was a protein that transports ornithine in the NCD-2 strain. Thus, it was hypothesized that Gms1961 functions as FenC and FenE. The analysis was performed using the Gms1960 protein and it had the greatest similarity with FenD (Additional File 1,Table S3); however, the FenD domains in Gms1960 and FZB42 varied greatly. Therefore, it was hypothesized that Gms1960 or other enzymes may have functions similar to those of FenD.
Although the fengycin synthetic gene cluster in the NCD-2 strain lacked two important genes-fenC and fenD-hat synthesize enzymes compared with the reported fengycin synthetic gene cluster, the NCD-2 strain was capable of producing 26 homologs of 5 kinds of fengycins. The amino acids at position 6 and 10 of the fengycin cyclic peptide ring determine the type of fengycin. There are currently five types of reported fengycins, A, B, A2, B2, and C (Additional File 1,Fig. S4). When the amino acid at position 6 was valine and at position 10 was isoleucine or valine, then fengycin B or fengycin B2, respectively, was produced (Fig. 5a, b) and (Additional File 1, Fig. S4); however, if the amino acid at position 6 was alanine, then fengycin A or fengycin A2, respectively, was produced (Fig. 5c, d) and (Additional File 1, Fig. S4). When the amino acid at position 6 was isoleucine or leucine and at position 10 was valine, then fengycin C was produced (Fig. 5e) and (Additional File 1, Fig. S4). The MS analysis of the fengycins in the NCD-2 strain revealed that the strain was capable of producing these five kinds of fengycins. Based on differences in the number of carbon atoms in the β-OH FA, fengycin had different homologs, and the molecular weight of each homologs differed by 14 (-CH2) [53]. The molecular structure of the lipopeptide determines its biological activity, and long-chain fatty acids increase the hydrophobic activities of lipopeptides, making them more likely to have membrane-bound antimicrobial effects [54]. A Bacillus circulans strain produces four fengycin homologs, but only fengycins with C16 and C17 carbon atoms in their β-OH FA chains had antibacterial activities [55]. The NCD-2 strain produced 14 fengycin homologs having more than 16 carbon atoms, and they accounted for a large proportion of all the homologs. It was speculated that these long-chain fengycins play important roles in the antimicrobial functions of NCD-2. The Bacillus siamensis SCSIO 05746 strain produces a great number of fengycin homologs, including 19 homologs of fengycin B [56]. Using an MS analysis, the five fengycins produced by the NCD-2 strain were divided into 26 homologs (Fig. 5a–e) and (Additional File 1, Fig. S5-S9). Therefore, NCD-2 is currently the strain with the largest number of known fengycin homologs [57].
During the microbial synthesis of secondary metabolites, such as lipopeptide, the relatively high energy-consuming process of protein synthesis takes priority [58]. Excessive energy consumption is not conducive to the normal growth of microbes, and, generally, microbes produce antibiotics in large amounts only when encountering pathogens or other stresses [59]. In the long-term evolution of NCD-2, the key synthetic genes fenEAB involved in synthesizing fengycin were conserved, while two important synthetic genes fenCD were lost. However, five fengycins are still produced. Gms 1961 played the dual roles of FenC and FenE, indicating that NCD-2’s fengycin synthetic process, which is unique to the strain, was more energy-efficient than the process used in the other strains..
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}