Infection can affect the development of atherosclerosis through two mechanisms: firstly, by directly infecting cells in the vessel wall, which increases the number of sites susceptible to lesions, and secondly, by indirectly infecting sites away from the atherosclerotic plaque, thereby activating the immune system which in turn increases the systemic inflammatory state.19 Poor oral hygiene has been recognized as an established risk marker for CVD.20 Studies have demonstrated that increased expression of CAP-mediated cytokines, including IL-2 and IL-6, leads to an initial inflammatory response in the rat aorta.21,22 Furthermore, studies have identified irreversible changes in the liver, indicating that CAP has systemic effects on multiple organs.22 Previous experiments have revealed that CAP promotes atherosclerosis in apoE−/− mice while altering the composition and diversity of the gut microbiota.10 Notably, fecal microbiota transplantation with altered gut microbiota as an independent variable, also exacerbated the degree of atherosclerosis in apoE−/− mice, suggesting a causal link between gut microbiota and atherosclerosis.23 Nevertheless, the precise mechanisms through which gut microbiota contribute to the development of atherosclerosis require further exploration. Based on this study's findings, it is possible that CAP impacts atherosclerosis by modulating the gut microbiota metabolites and intestinal permeability.
First, the Oil Red-O staining of the aortic root and aortic arch was used to evaluate the extent of atherosclerosis. Oil Red-O staining of frozen sections of aortic roots from 90 µm to 790 µm was used to evaluate plaque size (Fig. 1.B), and the image of the typical Oil-Red O staining in the aortic arch of the two groups is shown in the Fig. 1.C-D. The results show a statistically significant increase in aortic arch in the CAP group (Fig. 1.E, P < .01), suggesting that CAP causes an increase in the area of arterial endothelial injury in apoE−/− mice. The results showed that the atherosclerotic plaque volume at the aortic sinus was larger in the CAP group of mice (Fig. 1.F-G), CAP may have increased the extent of lipid accumulation in plaques. These findings are consistent with the results of the CAP model in apoE−/− mice fed with chow diet.10 Even though there are some significant differences in this modelling compared to previous CAP models. Firstly, the diet was different; this study used a high-fat diet instead of the previous chow diet. Secondly, to simulate of the clinical situation and refer to the modeling of multiple infections in experimental periodontitis,24 multi-infection construction CAP models were used. The results suggest that regardless of whether the mice are fed with a chow diet or an HFD, CAP can exacerbate atherosclerosis in apoE−/− mice.
HFD has been shown to affect the diversity and composition of the gut microbiota in humans and rodents,25 considering that HFD can alter the composition of gut microbiota, this study aimed to investigate whether CAP affects the composition and diversity of gut microbiota in apoE−/− mice fed with HFD. To assess the effects of endodontic infection with P. gingivalis on the gut microbiota composition and diversity while fed with an HFD, we performed 16S rRNA sequencing and analyzed the data using various bioinformatics tools. At the genus level, the top 20 altered taxa are shown (Fig. 2.A), and for display convenience we have also selected eight of them for presentation via box plots (Fig. 2.B-I). LEfSe was applied to the microbiota data of the CAP group and Con group, and four differentially abundant taxa were identified at the genus level (Fig. 2.J). Erysipelotrichaceae and Odoribacter were found to be increased, whilst Faecalibacterium and Lachnospiraceae were reduced in the CAP group compared to the Con group (Fig. 2.J). This result is consistent with clinical studies in which Erysipelotrichaceae was significantly increased in the gut microbiota of patients with atherosclerotic cardiovascular disease, heart failure syndrome with reduced ejection fraction, and heart failure syndrome with preserved ejection fraction.26 On the other hand, a decrease in Faecalibacterium and an increase of Lactobacillus were found in the gut microbiota of patients with clinical manifestations of atherosclerotic plaques with ischemic stroke,27 and the abundance of Faecalibacterium was negatively correlated with different markers of inflammation such as high sensitivity C-reactive protein and IL-6.28 The results showed that CAP increased the bacterial genera positively associated with atherosclerosis while decreasing those negatively associated. In terms of beta diversity, the gut microbiota of the CAP and Con groups were clearly distinguishable (Fig. 3.A, P = 0.007 in the permanova test). This finding suggests that CAP can still have significant effects on the composition of gut microbiota even fed with HFD. In terms of alpha diversity, Chao1 and Observed OTU indices represent the number of species in the microbiota, and Shannon and Simpson indices the diversity of the microbiota (Fig. 3.K). There was a reduction in the diversity and abundance of gut microbiota after CAP treatment with fed with HFD, but the differences were not statistically significant (Fig. 3.L-O, P > .05). Alpha diversity was significantly reduced in apoE−/− mice fed with HFD,29 considering the earlier findings 10, there was a relative increase in the diversity and abundance of gut microbiota after CAP treatment while fed with chow diet. The most likely cause of the alteration of alpha diversity is the combination of the effect of CAP and HFD, and the effect of HFD on alpha diversity was stronger than CAP. The most direct and commonly used research method to prove causality is FMT.30 Involvement of gut microbiota in the promotion of atherosclerosis by CAP was verified by FMT experiments in pseudo-sterile mice gavaged with faeces from CAP group mice (Supplementary data 1). It should be noted that the resolution and accuracy of 16S rRNA sequencing may be limited, and additional techniques such as shotgun metagenomics or metatranscriptomics may provide a more comprehensive characterization of the gut microbiota. In summary, CAP may influence the process of atherosclerosis by increasing harmful bacteria and decreasing beneficial bacteria.
Metabolites produced by gut microbiota play a crucial role in communication between the host and microbiota. The development of targeted and untargeted metabolomics has provided strong evidence that metabolites, such as short-chain fatty acids (SCFAs), bile acids, and trimethylamine N-oxide (TMAO), are involved in atherosclerosis pathogenesis.12–14 Targeted metabolomics measures a specific group of related metabolites with high precision, while untargeted metabolomics assesses a wide range of metabolites. Targeted metabolomics has limitations, including the inability to provide a comprehensive overview of the response and the inability to identify novel metabolites that may be relevant.31 Unlike targeted metabolomics, which is based on a pre-existing hypothesis, untargeted metabolomics can provide a broader and more comprehensive view of the metabolites involved in the link between CAP and atherosclerosis. Therefore, a combination of untargeted and targeted metabolomics was chosen for this study, with initial screening of the gut flora using untargeted metabolomics, followed by targeted metabolomics after key metabolites were found, in order to more comprehensively assess the accuracy of the metabolites involved in the link between CAP and atherosclerosis.
In the untargeted metabolomics analysis, all samples involved in the OPLS-DA analysis were within the 95% confidence interval region of the ellipse, and the results showed a strong separation effect between the two groups (Fig. 3.A, n = 10, P ≤ .05). The results indicate that CAP is associated with an alteration in the composition of metabolite in apoE−/− mice. The biological role of each metabolite was obtained by annotating them with the KEGG database br08001. The percentage content of each biological role was then counted and presented as a stacked histogram, the results showed an increase in metabolites related to lipid metabolism (Fig. 3.B), which has been established that disorders of lipid metabolism play an essential role in the pathogenesis of atherosclerosis.32 The magnitude of metabolite alterations and some metabolites of interest were determined by calculating the multiple of metabolite fold change and p-value (Fig. 3.D). TCDCA and cymarin were increased, while glucosamine 6-sulfate and phloretin were reduced in the CAP group. The alteration of TCDCA observed in this study is similar to the effects of aging on gut microbiota. High levels of serum TCDCA induced by aging activate TMAO-associated signaling pathways via hepatic FXR.33 Previous experiments also found increased serum levels of TMAO in mice from CAP group,23 suggesting that CAP may influence the development of atherosclerosis by modulating the gut microbiota metabolism of bile acids and affecting serum TMAO levels. This finding is consistent with metabolite set enrichment, which revealed an alteration in primary bile acid synthesis in the CAP group (Fig. 3.C). The CAP group showed high expression levels of primary conjugated bile acids, including TCDCA and taurocholate acid. For the function of glucosamine 6-sulfate, its ability to reduce insulin resistance and pro-inflammatory effects via combining with amyloid-β.34 Phloretin, a dihydrochalcone flavonoid, was found to be reduced in the CAP group (Fig. 3.D), which has been shown to have a thrombogenic and atherosclerotic preventive effect by inhibiting thrombin-mediated leukocyte-platelet-endothelial interactions,35 and low doses of phloretin have also been found to optimize lipid metabolism and prevent the acceleration of atherosclerosis in streptozotocin-induced diabetic mice.36 The purpose of this study was to investigate the correlation between alterations in the gut microbiota and its metabolites induced by CAP and the development of atherosclerosis. The percentage of lesion area and the nine candidate metabolites obtained by univariate analysis were used as environmental factors. These factors were used to perform correlation clustering heat map analysis with CAP-induced alterations in the gut microbiota at the genus level (Fig. 3.E). The red and blue colors in the graph represent the intensity of correlation from high to low. The percentage of lesion area was found to cluster with TCDCA and TCDCA sodium salt. Based on correlation and cluster analysis, indicates that the gut microbiota and its metabolites have similar patterns, which is consistent with our assumptions. The gut microbiota of the CAP group showed enrichment of Erysipelotrichaceae, Lachobacillus and Staphylococcus (Fig. 2. D, E, I). These genera were found to have a positive correlation with the severity of atherosclerosis or deleterious metabolites. The TCDCA was found to be positively correlated with Lactobacillus (Fig. 3.E), and it was reported to be associated with increased systemic inflammation and intestinal permeability caused by a ketogenic diet.37 Additionally, another research also showed an increased level of primary taurine conjugated bile acids, including taurocholic acid and TCDCA, in mice treated with Lactobacillus casei Strain38. Notably, Staphylococcus was positively associated with the extent of atherosclerosis, this result is consistent with research showing that Staphylococcus was enriched in four thrombus samples taken from patients undergoing thrombectomy.39 In contrast, a positive correlation was found between beneficial bacteria and beneficial metabolites. For instance, Faecalibacterium was positive correlation with glucosamine 6-sulfate, which is the functional group of enoxaparin responsible for the anti-inflammatory effect.40 There was also a negative correlation between beneficial metabolites and enriched genus in the CAP group. Of the more significant ones, Odoribacter showed a negative correlation with both phloretin and glucosamine 6-sulfate (Fig. 3.E). In summary, CAP leads to a decrease in beneficial gut microbiota and metabolites and an increase in deleterious gut microbiota and metabolites. These alterations are positively correlated with atherosclerosis.
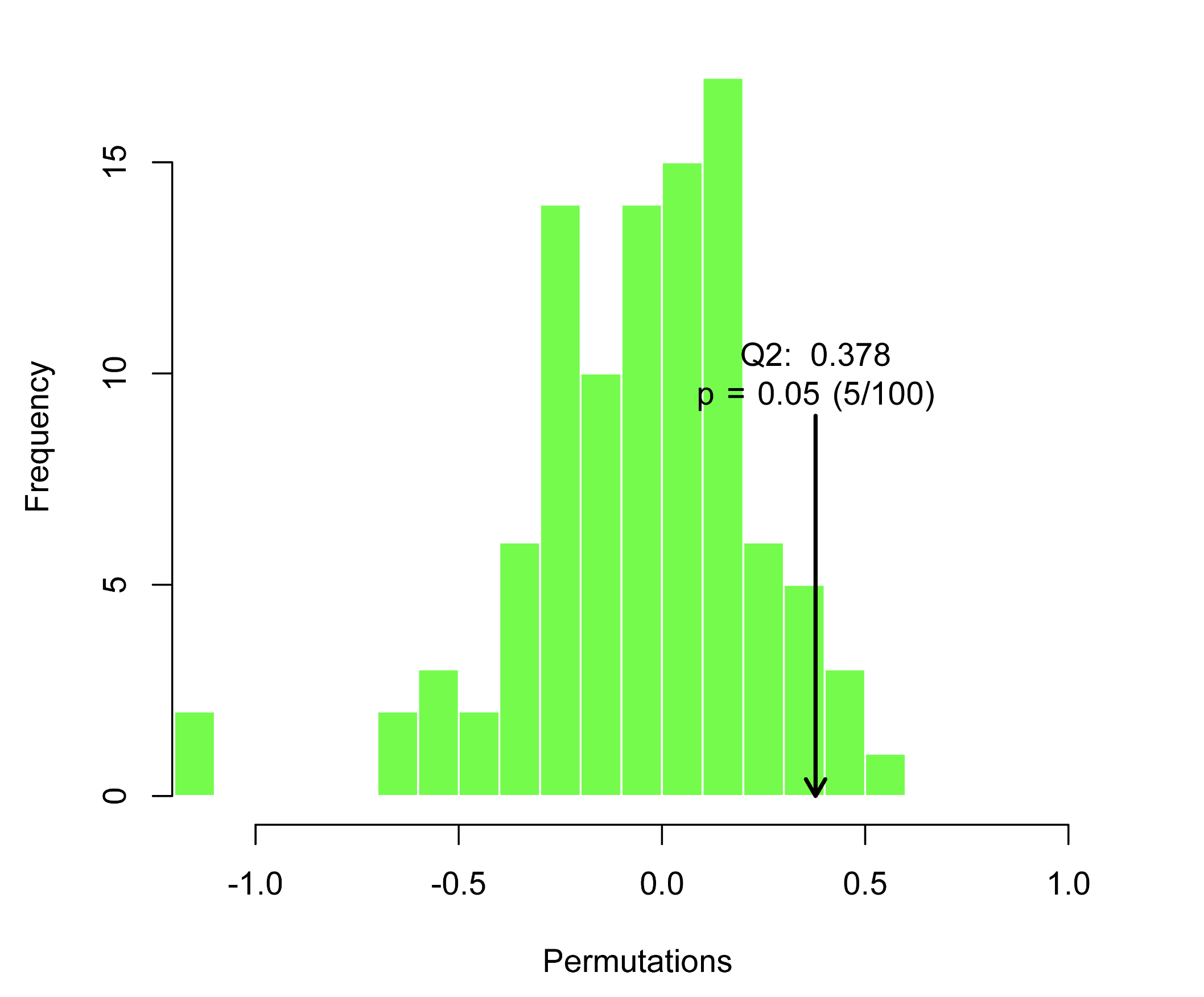
Primary bile acid synthesis may play a crucial role in the promotion of atherosclerosis by CAP through non-target metabolomics data. Further, the faeces were examined for bile acid metabolomics, OPLS-DA analysis of the composition of bile acids in the CAP group was clearly distinguished from that of the Con group (Fig. 4.A), and the difference was statistically significant after Permutation testing (Supplementary data 3, P < .01, Q2 = 0.767), Subsequent screening for differential bile acids showed increased expression of TCDCA, TCA, TDCA and decreased expression of UCA, 6,7-diketoLCA in the CAP group (Figure B-C). The use of Obeticholic acid was found to prevent the development of atherosclerosis by inhibiting intestinal cholesterol absorption while TCA levels were found to be reduced by 71%.41 Carotid intima-media thickness (C-IMT), a proxy for subclinical atherosclerosis, was analysed in correlation with patients' serum metabolic profiles, it was found that deoxycholic acid and taurodeoxycholic acid (TDCA) levels were significantly higher in patients with abnormal C-IMT than in those with normal C-IMT.42 The levels of TCDCA, TCA were increased in gingival tissues of patients with periodontitis and showed a positive correlation with periodontal clinical parameters.43 These results suggest that CAP-induced changes in bile acids may contribute to the extent of atherosclerosis by affecting intestinal permeability.
The intestinal barrier forms a barrier between the intestine and the external environment, regulating intestinal contents such as bacteria and the immune system interactions. mucin is crucial in the prevention of intestinal barrier, and Muc-2 is a key component of mucin in the colon, produced by goblet cells, which form an internal and external mucus layer and cover the epithelial cell surface.44 Whereas tight junction protein, such as ZO-1, occludin, and claudin, is an important part of the intestinal epithelial barrier, which regulates the permeability of the intestinal epithelial barrier and prevents intestinal pathogenesis, increased permeability of tight junction proteins when damaged.45 Considering that bile acids may promote atherosclerosis by increasing intestinal permeability through disruption of the intestinal barrier, we stained tissue sections of intestines and showed that decreased villus height, decreased intestinal mucus expression and intestinal mucin expression (Fig. 5.A), and decreased intestinal tight junction protein in the intestine (Fig. 6.A-B), suggesting that CAP causes damage to the intestinal mucus barrier and the intestinal mechanical barrier. Activation of the ERK 1/2 signalling pathway is involved in mediating the down-regulation of intestinal tight junction proteins,46 and LPS stimulation of intestinal epithelial cells also revealed activation of ERK 1/2 leading to a decrease in tight junction proteins.47 This is consistent with our results that the ERK1/2 signalling pathway was found to be activated in colonic tissue. The FD-4 in vivo assay was used to examine intestinal permeability in mice, and the results confirmed that CAP promotes intestinal permeability (Fig. 6.C), and a correlation analysis between the degree of atherosclerosis and permeability in mice showed a strong positive correlation between the two (Supplementary data 4.C, P < .05, correlation coefficient = .564). Consistent with our results, impaired intestinal barrier function can result in bacterial translocation and the presence of bacterial products in the circulation, potentially leading to atherosclerosis and chronic heart failure.48 Our results suggest that CAP may promote the development of atherosclerosis by causing increased intestinal permeability.
In summary, this study unravels the complex interplay between oral health, gut microbiota, metabolites, and cardiovascular disease (Fig. 7). It underscores the significance of maintaining oral hygiene and treating chronic apical periodontitis to promote cardiovascular well-being. Moreover, the findings offer valuable insights into potential therapeutic strategies aimed at modulating gut microbiota and metabolic pathways to mitigate the impact of CAP on atherosclerosis development. Overall, this research contributes to our understanding of the multifaceted nature of cardiovascular diseases and offers prospects for novel interventions.
{kind=link}
{kind=link}
{kind=link}