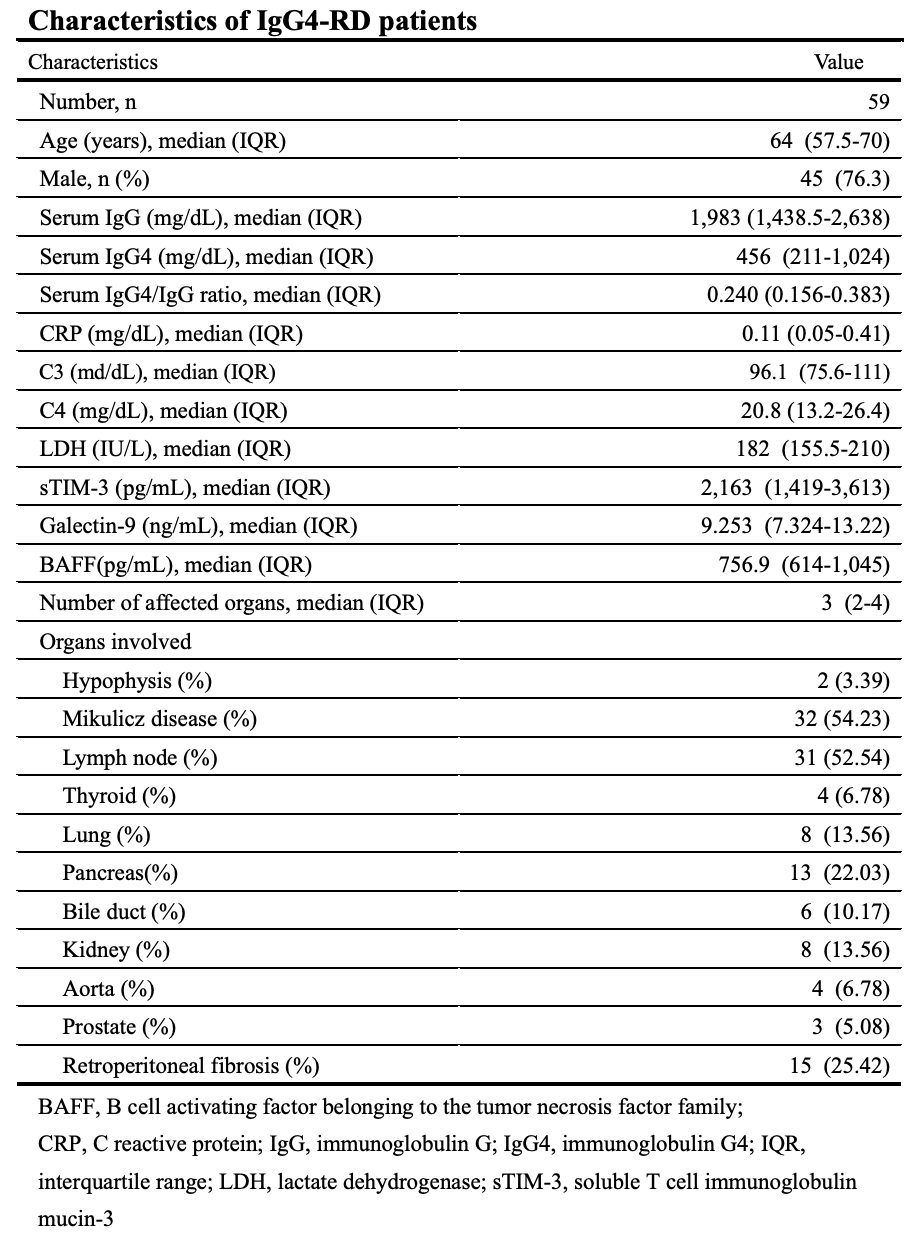
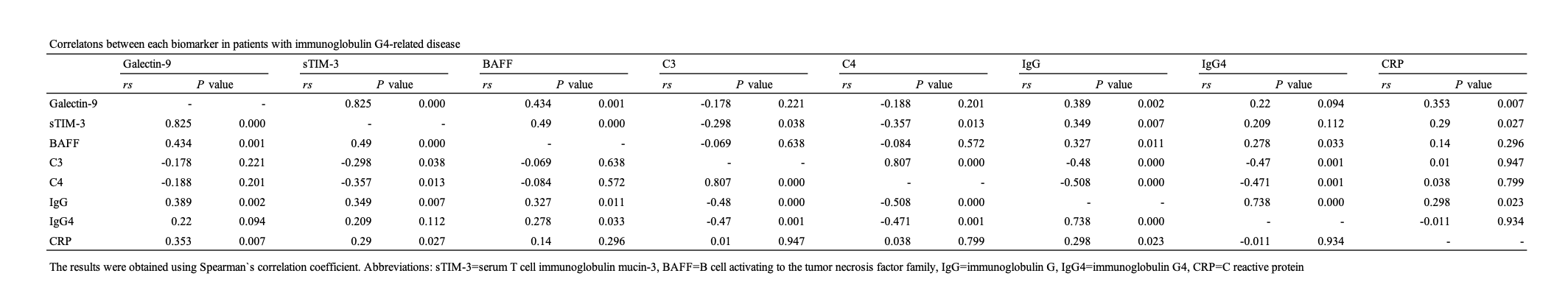
Galectins, a protein from a family of lectins with affinity for β-galactoside-containing oligosaccharides, are expressed by the immune cells (22). Recent studies have shown that galectins play crucial regulatory roles in inflammationtand autoimmunity (23). In this study, we evaluated the serum levels of sTIM-3 and its ligand molecule, Gal-9, in IgG4-RD patients. Although circulating Gal-9 or sTIM-3 were not correlated with the serum IgG4 levels or the ratio of IgG4/IgG, our results indicated that the serum levels of Gal-9 are significantly elevated in patients with IgG4-RD as compared to those in HCs and RA patients. Furthermore, the serum levels of sTIM-3 are significantly elevated in patients with IgG4-RD as compared to those in HCs. These findings indicate that these checkpoint molecules could be involved in the pathophysiology of IgG4-RD.
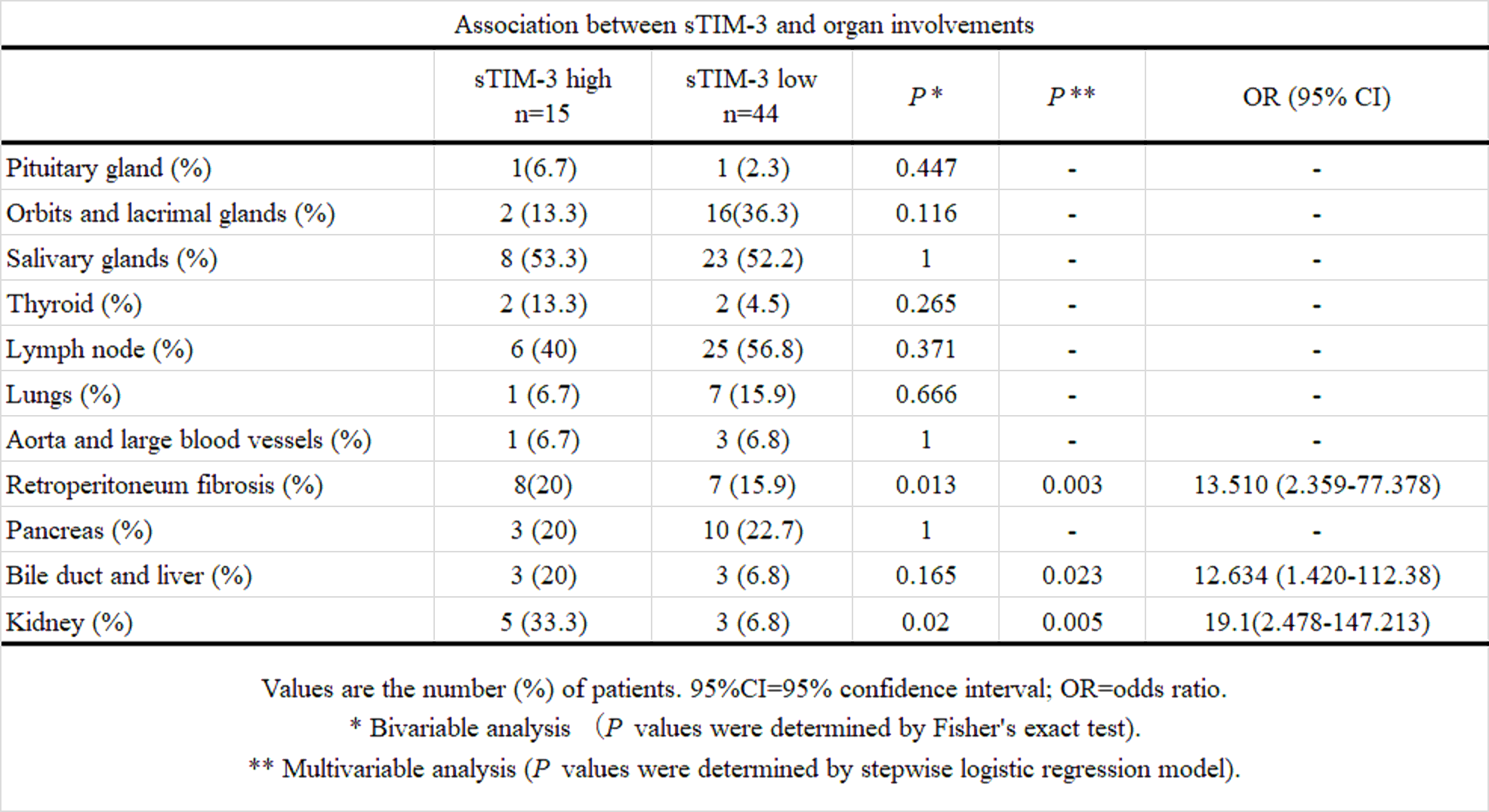
Previous studies have shown the upregulations of Th2 cytokines (interleukin (IL)-4, IL-5, IL-13 and IL-21) and the regulatory T cell-mediated cytokines (IL-10 and transforming growth factor-β) in IgG4-RD patients (7). Therefore, IgG4-RD appears to be driven by pathogenic Th2 cells or a combination of Th2 cells and regulatory T cells (Treg cells), and these T cell subsets may activate macrophages and fibroblasts that cause inflammatory and fibrotic processes in the affected organs (24). We hypothesized that Th1/Th2 imbalance in IgG4-RD is associated with the dysregulation of checkpoint molecules and analyzed their associations with the clinical phenotypes of IgG4-RD. In contrast to patients with organ involvement limited to the lacrimal or salivary glands, patients with visceral involvements presented with higher levels of serum sTIM-3. Furthermore, we observe an organ-specific increment in sTIM-3, particularly biliary, kidney, or retroperitoneum involvement in IgG4-RD. Our data indicated that elevated levels of sTIM-3 in patients with IgG4-RD could be related to the clinical phenotypes of IgG4-RD, including its patterns of organ involvements.
TIM-3 acts as a co-inhibitory receptor that is expressed on exhausted T cells. TIM-3 was initially thought to be expressed only by T cells; it has now been proven that TIM-3 is expressed by multiple cell types, including DCs, macrophages, and Tregs (25), indicating that TIM-3 also functions as an inhibitory receptor in these cells (26). Gal-9 has been identified as a ligand for TIM-3 (27); however, its putative ligands other than Gal-9 and its inhibitory effect on T cell remain unclear. The precise cellular interaction within sTIM-3 and its ligands remains unclear; however, it is possible that a combination of sTIM-3 with its ligands competitively reduces the inhibitory effect on the pathway following TIM-3 on immune cells (28), resulting in Th1/Th2 cell imbalance in IgG4-RD (29). The association between th2 immune response and M2 macrophage play an important role in the pathogenesis of IgG4-RD (30). Furthermore, infiltration of M2 macrophage to affected organ in IgG4-RD may promote fibrosis (31). Because Gal-9/TIM-3 signaling pathway inhibit M1 macrophage polarization by short-term lipopolysaccharide stimulation, M2 macrophage is relatively dominant (32). Our result showed sTIM-3 was increased in IgG4-RD patients. However, the number of TIM-3 on macrophage was not evaluated. The expression of TIM-3 on macrophage may be upregulated in IgG4-RD patients. Gal-9/TIM-3 signaling pathway may cause fibrosis in affected organs by promoting the differentiation of M2 macrophages.
Recent studies have identified several new immune checkpoint targets, such as lymphocyte activation gene-3 or TIM-3. The investigations on these molecules have generated new findings in the pathophysiology of immune-mediated disorders (33). Tfh is critical regulator of immune responses in inflammatory disorders (34–36). Checkpoint inhibitors are known to induce B-cell activation and class-switched IgG production; these processes are dependent on the Tfh cell function (20). Furthermore, the case of an IgG4-RD patient with lung cancer receiving checkpoint inhibitors has been reported (18). These findings suggest that the interaction between B cells and Tfh cells through these checkpoint molecules could be involved in IgG4 class switching (18). Tfh could be subdivided to distinctive functional subsets as per the checkpoint molecules expressions. programmed cell death 1 demarcated potent Tfh subsets (37), and TIM-3 appears to be associated with reduced Tfh function (38), suggesting that the co-inhibitory Gal-9/TIM-3 pathway can limit the functions of Tfh (39). Potent Tfh might play a crucial role in IgG4 class switching (40); therefore, elevated levels of sTIM-3 may affect the Tfh functions by interfering with the Gal-9/TIM-3 co-inhibitory checkpoint systems. These co-inhibitory checkpoint pathways may alter the interaction between Tfh cells and plasmablasts (41) in the immunopathology of IgG4-RD and its expanding organ involvement. Given the multi-organ nature of IgG4-RD, the predictors for particular organ involvements would be valuable for clinical application and research of IgG4-RD.
Although the serum concentrations of sTIM-3 were associated with visceral involvements of IgG4-RD, the levels were not modulated by steroid treatment. Steroid is effective for most patients with IgG4-RD (2), however, the relapses of IgG4-RD were frequently observed during the steroid tapering periods (2). Steroid is considered to be first-line treatment for remission induction in IgG4-RD (42), however, it is possible that the incomplete regulation of immunopathology of IgG4-RD could be linked to this increased serum levels of immuno-checkpoint molecules even after the steroid treatment.
Additional studies are required to clarify the roles of circulating checkpoint molecules in the pathophysiology of IgG4-RD, and their relationship to the clinical phenotypes. Furthermore, it is necessary to elucidate whether more aggressive treatment such as B cell-depletion treatment may affect the upregulated immune-checkpoint molecules in IgG4-RD.
There are certain limitations of this study. The sample size was relatively small, and larger scale studies are necessary to confirm the present finding. Checkpoint molecules profiles in other potential controls, such as those with Sjögren's syndrome or lymphoproliferative disorders, were not compared with the profiles in IgG4-RD patients in the present study. Because of inadequate number of untreated RA patients, more than half of the control patients of RA was already treated. This study was a cross-sectional analysis for untreated IgG4-RD patients. Therefore, clinical manifestations that occurred during the disease course were not completely surveyed. We enrolled only Japanese patients with IgG4-RD; therefore, non-Japanese patients with IgG4 were not included, and further studies on non-Japanese patients with IgG4-RD are warranted. The mechanism through which the Gal-9/TIM-3 pathway contributes to the pathogenesis of IgG4 was not clarified because the functional analysis by in vitro cannot be performed.
In conclusion, we demonstrated that serum sTIM-3 and Gal-9 levels were elevated in IgG4-RD patients. With respect to the relationship with the clinical phenotypes, the serum sTIM-3 levels were significantly higher in patients with visceral involvements than in those with disease limited to the lacrimal or salivary glands. These data suggest that the circulating checkpoint molecules are involved in the pathophysiology of IgG4-RD and their relationship to its patterns of organ involvement.
{kind=link}
{kind=link}
{kind=link}