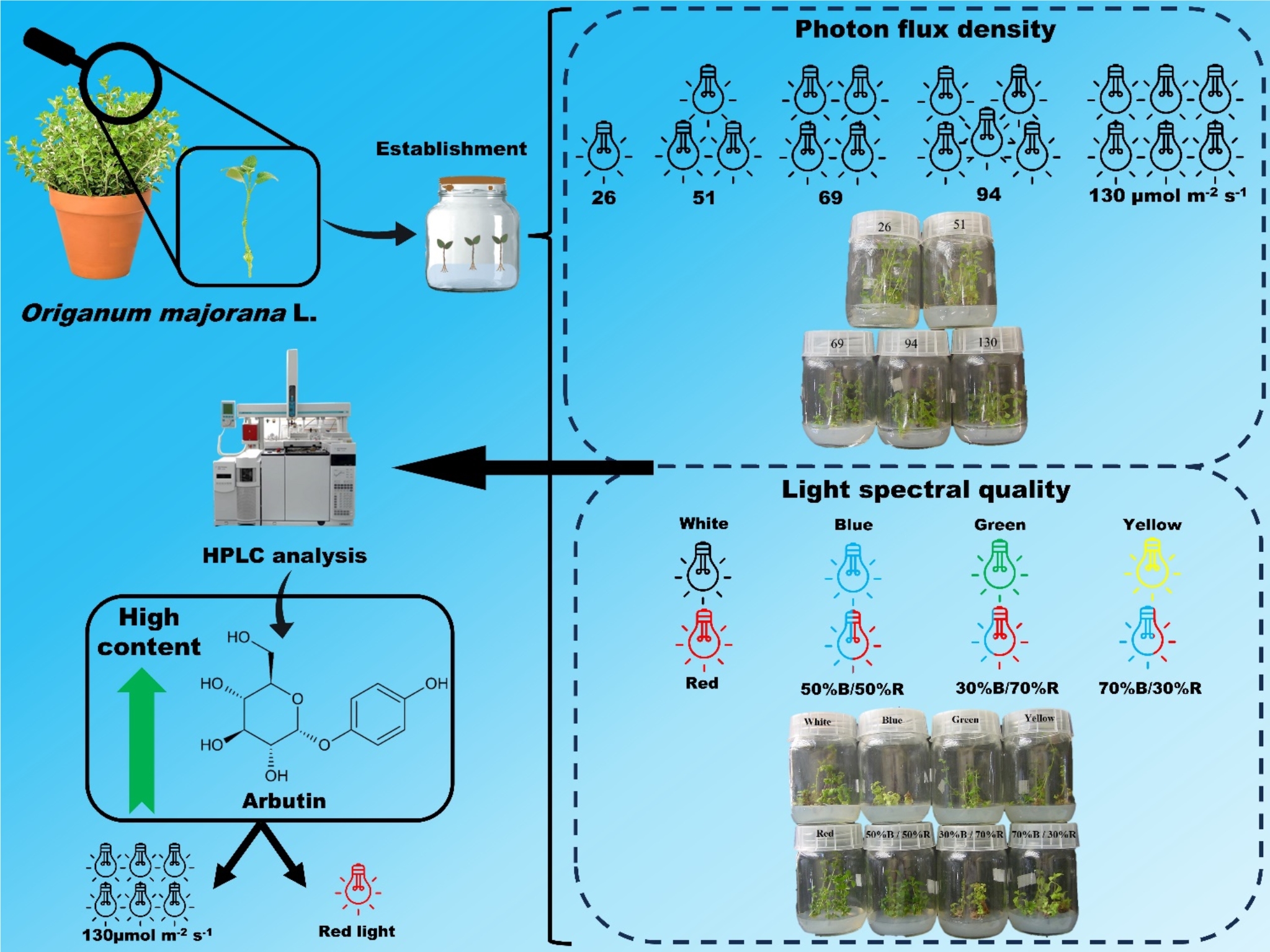
Analysis of growth under light intensity
The in vitro growth of O. majorana was affected by different light intensities. Root system induction in O. majorana plantlets was very low, so there was no condition to compute this parameter. Etiolation was observed in plantlets at 26 to 51 µmol m-2 s-1 with thin stems and smaller leaves compared to treatments 94 and 130 µmol m-2 s-1 (Fig. 1). On average, the highest number of shoots (2.6) and shoot length (6 cm) were observed under cultivation at 51 and 94 µmol m-2 s-1, respectively. The in vitro cultivation of O. majorana under increased light intensity tended to reduce the number of regenerated shoots (Fig. 2a). In H. suaveolens cultivation, increased light intensity (13 to 69 µmol m -2 s -1) increased the number of shoots and shoot length, with the best growth being observed under light intensities of 47 and 69 µmol m -2 s -1 (Andrade et al. 2017). In the in vitro culture of Withania somnifera, the plantlets had higher growth under cultivation at 30 µmol m-2 s-1 (Lee et al., 2007). Higher light intensities were favorable for leaf and stem expansion of O. majorana, probably because light intensity regulates the size of leaves and stems, as well as their morphogenesis. Therefore, controlling light intensity, wavelength and photoperiod allows the production of plants with the best desired characteristics (Cavallaro et al. 2022). Many abiotic factors, such as light, can affect plant growth in vitro, thus making it difficult to choose a more effective light intensity and spectrum for many species. This shows that each genotype may respond differently according to the light regime subjected.
Different light intensities significantly influenced (p < 0.05) the production of leaf dry weight (LDW), stem dry weight (SDW) and aerial part dry weight (ShDW). A greater gain of stem and leaf dry weight was observed from 69 µmol m− 2 s− 1 (Fig. 2). The intensity of 94 µmol m− 2 s− 1 was higher than all other treatments regarding leaf dry weight (48.8 mg − 1), stem (34.7 mg − 1) and aerial part dry matter (83.5 mg − 1). The plantlets of U. dioica (Coelho et al. 2021)d gracilis (Lazzarini et al. 2018) cultivated under 94 µmol m− 2 s− 1 also obtained greater dry weight gain and growth.
This study and others confirm the importance of light intensity in growth and dry weight accumulation in plants grown in vitro. The results of this study and others showed that an intensity above 94 µmol m− 2 s− 1 affected the dry weight accumulation in the plantlets. The O. majorana plantlets at the highest intensity of 130 µmol m− 2 s− 1 reduced dry weight accumulation, which may have occurred due to the reduced efficiency of photosynthesis and damage to the reaction center (Fan et al. 2013; Hsie et al. 2019). When plantlets are under prolonged excessive irradiation, photoinhibition can occur, leading to impaired growth and development (Kitao et al. 2000). Other studies also reported a reduction in the total dry weight gain of plantlets grown in vitro under high light intensities, such as those of U. dioica (Coelho et al. 2021), L. rotundifolia (Hsie et al. 2019)d amboinicus (Silva et al. 2017).
The dry weight gain of plant organs varies with the species. The cultivation of Gerbera jamesonii with increasing light intensities from 40 to 120 µmol m− 2 s− 1 did not affect dry weight production or the number of shoots (Cioć et al. 2019). The in vitro cultivation of H. suaveolens under different light intensities showed an increase in growth variables with increasing light intensity from 13 to 69 µmol m− 2 s− 1 (Andrade et al. 2017). Other species respond to different intensities, for example, A. suaveolens (Araújo et al. 2021)d dulcis (Rocha et al. 2022) at light intensities of 139 µmol m− 2 s− 1, P. amboinicus 69 µmol m− 2 s− 1, and L. rotundifolia at low intensities of 20 to 54 µmol m− 2 s− 1 (Hsie et al. 2019).
The lowest values of dry weight were observed in plantlets grown at 26 µmol m− 2 s− 1 (Fig. 2). In general, plants grown in low light environments have lower biomass values due to the low production of photoassimilates, which is the result of inefficient photosynthesis. In the in vitro culture of A. millefolium, the highest values of shoot dry weight and area length were observed under cultivation at 27 µmol m− 2 s− 1 (Alvarenga et al. 2015).
Analysis of photosynthetic pigments under light intensity
The different light intensities significantly (p < 0.05) affected the concentration of photosynthetic pigments of O. majorana plantlets grown in vitro. The highest values for chlorophyll a and b and carotenoids were observed in cultivation under a light intensity of 26 µmol m-2 s-1. The species L. rotundifolia also obtained higher values of photosynthetic pigments under a low light intensity of 20 µmol m-2 s-1 (Hsie et al. 2019). In this study, the lowest values of chlorophyll and carotenoids were observed in cultivation under intensities of 94 and 130 µmol m-2 s-1 (Table 1).
Table 1
Concentration of photosynthetic pigments in Origanum majorana plantlets cultivated in vitro under different light intensities, at 40 days.
| Intensity µmol m− 2 s− 1 | Chlorophyll | | | Carotenoids |
| a | b | Total | |
| (mg/g FM) |
| 26 | 0.59a | 0.14a | 0.73a | 0.15a |
| 51 | 0.36c | 0.11b | 0.47c | 0.10c |
| 69 | 0.46b | 0.11b | 0.57b | 0.12b |
| 94 | 0.29d | 0.07d | 0.36d | 0.08d |
| 130 | 0.28e | 0.08c | 0.37d | 0.10c |
| CV (%) | 11.76 | 13.39 | 11.23 | 9.30 |
| Means followed by the same letter in the column do not differ from each other, according to the Scott Knott test, at the 5% probability level. FM: fresh matter. |
Light intensity is one of the key factors in the regulation of genes responsible for chlorophyll and carotenoid biosynthesis. The results obtained in the present study indicate that the values of chlorophyll and carotenoids decrease in O. majorana with increasing light intensity, thus suggesting a correlation between greater light intensity and reduced concentration of photosynthetic pigments due to photodegradation (Cavallaro et al. 2022; Silva et al. 2017). The plantlets are grown in flasks where there is high humidity, resulting in less epicuticular wax (EW) on the leaves. EW has been identified as an important physical adaptation to a wide range of environmental factors, such as droughts, high irradiance and herbivory. Thus, in vitro plantlets with lower wax accumulation become susceptible to high light conditions and prone to photoinhibition due to damage to the photosynthetic apparatus and photosynthetic pigment synthesis (Silva et al. 2017; Singh and Patel 2014). Nevertheless, in this study, dry weight accumulation was not affected by high radiation in O. majorana (Fig. 2).
However, not all species exhibit this behavior in vitro. In the cultivation of A. millefolium, when evaluating different light intensities (13 to 69 µmol m− 2 s− 1), the authors observed a higher content of photosynthetic pigments at 69 µmol m− 2 s− 1 (Alvarenga et al. 2015). Similar results were described in the in vitro cultivation of G. jamesonii under different light intensities (40 to 120 µmol m− 2 s− 1), where the photosynthetic pigment content increased with increasing intensity, with an intensity of 80 µmol m− 2 s− 1 accumulating higher pigment contents (Cioć et al. 2019). Thus, it is clear that different species in vitro need different light conditions, adapting morphologically and physiologically to different light intensities in a unique way.
Quantification of arbutin under different light intensities
Light is an abiotic factor that affects the secondary metabolite compounds of plants. In this study, different light intensities significantly affected arbutin accumulation in Origanum majorana plantlets grown in vitro (Figs. 3 and 4). The increase in light intensity stimulates the production of phenolic compounds in plants, with the function of protecting the photosynthetic apparatus, because light affects the production of secondary metabolites such as phenolic compounds and flavonoids, however, its action in increasing or decreasing the content of compounds is specific to the species and the constituent of interest (Jing et al. 2018; Taulavuori et al. 2018).
The increase in light intensity promoted greater accumulation of arbutin in the leaves of O. majorana in vitro. From the light intensity of 51 µmol m− 2 s− 1, arbutin accumulated more than twice compared to the control (26 µmol m− 2 s− 1). The increase in light intensity leads to a greater production of photoassimilates to a level where harmful reactions to the plant begin to occur, caused by light stress (Taiz et al. 2017). Light stress did not favor the accumulation of photosynthetic pigments, as discussed above, but favored the accumulation of arbutin (12.5 mg/g) as a possible response to light stress in plantlets grown under 139 µmol m− 2 s− 1. Quantitative analysis of Arbutus andrachne L. leaves in in vivo and in vitro cultivation using high-performance liquid chromatography (HPLC) showed that the in vivo leaves contained a higher arbutin content (0.3–0.81% w/w) than in vitro leaves (0.09% w/w) (Mostafa et al. 2010). In this study, in vitro cultivation of O. majorana using HPLC revealed that the leaves contained an arbutin content of 1.25% under the highest light intensity.
Similar results with the secondary metabolites regarding light irradiance were observed in the in vitro culture of Digitalis mariana under different light intensities (20 to 139 µmol m− 2 s− 1), where the increase in light intensity led to an increase in the cardenolide content, justifying this increased stress due to exposure to high light intensity (Silva et al. 2022). Another study with in vitro cultivation under different light intensities changed the profile of volatile compounds and increased the content of carvacrol in P. amboinicus (Silva et al. 2017). In contrast to the in vitro cultivation of L. gracilis under different light intensities, researchers observed an increase in the y-terpinene content and a decrease in the carvacrol content under cultivation at 26 µmol m− 2 s− 1 (Lazzarini et al. 2018).
Growth analysis under different wavelengths
The wavelength can affect secondary metabolism, shoot multiplication and the development and growth characteristics of the planlets in vitro, i.e., shoot length, dry weight and photosynthetic pigments. The growth of O. majorana in vitro was significantly influenced by the spectral quality of light (Figs. 5 and 6). The number of shoots regenerated in most different light spectra had no significant effect (Fig. 6A). Plantlets grown under green and yellow light were etiolated and had small leaves. Higher values of shoot length were observed under cultivation under the white light spectrum (7.3 cm). This result can be explained because white light has a broad spectrum, which is different from the specificity of monochromatic lights (Cavallaro et al. 2022). However, monochromatic blue light spectra and combinations of blue and red light had an inhibitory effect on shoot length (Fig. 6B). Different aspects of plant growth and development are regulated by various blue light receptors. The wavelength region of light from UV-A to blue (B, 320–500 nm) is perceived by three photoreceptors that act in plants. The three photoreceptors act as chromophores called cryptochromes (CRY1, CRY2, and CRY3). At high blue light irradiation, CRY1 inhibits stem growth and reduces internode elongation, and CRY2 inhibits it at low blue light irradiation (Cavallaro et al. 2022; Lin 2000).
In cultivation under monochromatic red light and a combination of blue (50%) and red (50%) light, the plantlets were visually more vigorous, with larger leaves and dark green color (Fig. 5). The leaf dry weight (LDW) of plantlets grown under white light accumulated the highest value (28.1 mg), followed by plantlets grown under a combination of blue and red light (70% B:30% R) (Fig. 6C). Plantlets grown under monochromatic blue, green and yellow light accumulated less dry weight (Fig. 6, C, D, E). White- and red-light spectra were more efficient in the production of SDW, with blue and green monochromatic lights, and 50% B:50% R showed lower accumulation of SDW. In general, higher aerial part dry weight (ShDW) production was favored by cultivation in the white- and red-light spectra and a combination of blue and red (70% B:30% R and 30% B:70% R).
Different species responded to dry weight accumulation independently. The cultivation of U. dioica under the combination of 70% R:30% B resulted in higher dry weight accumulation than that under white light (Coelho et al. 2021). The species L. rotundifolia also obtained the best biometric data under the combination of 70% R:30% B (Hsie et al. 2019). The species A. suaveolens obtained the highest dry weight gain under white and fluorescent light (Araújo et al. 2021). For the light spectra, L. dulcis plantlets grown under combinations of 30% B:70% R and 50% B:50% R provided greater light accumulation of dry weight (Rocha et al. 2022). Monochromatic red led to a greater gain of dry weight in the species P. amboinicus (Silva et al. 2017) and in L. gracilis (Lazzarini et al. 2018). All these results indicate that the responses of plants to LED lighting are species and/or cultivar dependent.
Analysis of photosynthetic pigments under different wavelengths
The accumulation of photosynthetic pigments was significantly influenced by the light spectrum (Table 2). The leaves of O. majorana plantlets accumulated higher levels of chlorophyll a, total chlorophyll and carotenoids under cultivation under green light. There was a significant increase in chlorophyll a content under green light (0.68 mg/g FW), followed by the red-light spectrum (0.46 mg/g FW). High values of photosynthetic pigments observed under green light cultivation are indicative of plant stress, and this increase is a way to compensate for the lack of photosynthetically active light (Cavallaro et al. 2022). This compensation is explained by the penetration of green light into the plants, which favors plant growth, increasing photosynthesis in the leaves more efficiently than blue and red light (Alvarenga et al. 2015). Similar results were observed in the cultivation of A. millefolium under the green light spectrum, which showed higher values of chlorophyll a and b and carotenoids (Alvarenga et al. 2015).
Table 2
Concentration of photosynthetic pigments in Origanum majorana plantlets cultivated in vitro under different light spectra, at 40 days. FM: fresh matter.
| Spectra | Chlorophyll | | | Carotenoids |
| a | b | Total | |
| (mg /g FM) |
| White | 0.41c | 0.16b | 0.57b | 0.12c |
| Blue (B) | 0.23f | 0.06d | 0.29e | 0.07f |
| Green | 0.68a | 0.16b | 0.80a | 0.17a |
| Yellow | 0.16g | 0.04e | 0.20f | 0.05d |
| Red (R) | 0.46b | 0.11c | 0.57b | 0.13b |
| 50%B:50%R | 0.33d | 011c | 0.44d | 0.11d |
| 30%B:70%R | 0.29e | 0.22a | 0.50c | 0.10e |
| 70%B:30%R | 0.45b | 0.12c | 0.57b | 0.13b |
| Means followed by the same letter in the column do not differ from each other, according to the Scott Knott test, at the 5% probability level. FM: fresh matter. |
Lower accumulation of chlorophyll a and b was observed under the yellow (0.16 and 0.04 mg/g FW, respectively) and blue (0.23 and 0.06 mg/g FW) spectra (Table 2). Weremczuk-Jeżyna et al. (2021) observed contrary results, where the highest chlorophyll content in the cultivation of Dracocephalur forrestii was observed under the blue light spectrum. Likewise, in the cultivation of Saccharum officinarum, the combination of blue- and red-light spectra (70% B:30% R and 50% B:50% R) induced the production of total and chlorophyll a, the authors justify these results by the fact that blue light blue light plays an important role in the synthesis of chlorophyll, inducing its production and chloroplasts (Silva et al. 2014). In the in vitro culture of P. amboinicus, they observed an increase in the production of photosynthetic pigments under the spectra of the combinations of 50% B:50% R and 30% B:70% R (Silva et al. 2017). Therefore, according to data in the literature compared to the results of the present study, it was noted that the increase in photosynthetic pigments was dependent on species and light quality.
The spectral quality of light significantly influences the biological response of the aerial part. Since plant photoreceptors responsible for plant development and photosynthesis are known to be stimulated mainly and most significantly by the red and blue regions of the light spectrum, most studies have evaluated the impact of monochromatic red (660 nm), blue (460 nm) and combined blue (440–480 nm) with red lights (630–665 nm). Little information is available on the effects of the far red, green and yellow regions of the spectrum (Cavallaro et al. 2022).
Quantification of arbutin at different wavelengths
The arbutin levels in O. majorana plantlets were affected by the spectral quality of light (Fig. 7). The chromatographic profile of the plantlets grown under the different light spectra can be seen in Fig. 8. Higher values
of arbutin production were observed under cultivation under red monochromatic light (13 mg/g), 35% higher than under white light (9.6 mg/g) and 2.8 times higher than under monochromatic yellow light. The study shows that the plantlets that were cultivated with the highest proportion of red in relation to the blue spectrum (30% B:70% R) had the highest arbutin accumulation (10.3 mg/g). The red monochromatic light accumulated 94% more than the blue monochromatic light (6.7 mg/g). The use of monochromatic or composite LED lights with different spectral qualities may be a tool for the induction of bioactive metabolites and pigments of interest in in vitro culture. Lower values of arbutin were observed under blue (6.7 mg g− 1), green (6.5 mg g− 1) and yellow (4.6 mg g− 1) light culture. These results can be explained by the fact that plants respond to the environment differently, allocating their photoassimilates for the biosynthesis of elements necessary for their survival and adaptation to the environmental conditions offered by different light sources.
The reports that the variation in secondary compounds under cultivation at different wavelengths is species dependent. The in vitro culture of D. mariana cultivated under combined red and blue light (30% R:70% B and 50% R:50% B) showed higher cardenolide content (Silva et al. 2022). Red and blue light are mostly responsible for the biosynthesis of secondary metabolites due to their participation in photosynthesis. Silva et al. (2017) reported a higher accumulation of secondary metabolites in the in vitro culture of P. amboinicus under the blue light spectrum, where they obtained a higher carvacrol content. The monochromatic blue spectrum increased the content of myrcene and limonene compounds and the highest content of pentadecane under the white spectrum and combinations of red and blue in the essential oil of L. rotundifolia (Hsie et al. 2019). The volatile compounds in the species A. suaveolens increased the linalool content under white LED light (Araújo et al. 2021). The wavelength of 30% R:70% B obtained a 5.53-fold increase in the concentration of phenolic compounds and an 8.63-fold increase in the concentration of flavonoids compared to the fluorescent lamp (Coelho et al. 2021). Several studies emphasize that the production of phenolic compounds depends on the species, cultivar, and exposure time and that each phenolic compound has a specific response to light quality (Loi et al. 2021). Since arbutin is a phenol, its production in O. majorana under different light spectra is expected to be species specific.
Principal Component Analysis (PCA)
PCA (scores and loadings) was used to evaluate the correlations between dry weights, arbutin content and photosynthetic pigments of O. majorana treated under different wavelengths and light intensities. The PCA of light intensity was based on 8 parameters (growth and chemical compounds) that accounted for 96.51%, and for light quality, it was based on 6 parameters that accounted for 84.6% of the total variation in the experimental procedure. Based on these parameters, it was possible to group the data collected into 3 clusters to evaluate light quality (Fig. 9A) and intensity (Fig. 9B).
Evaluating the correlations of the 6 parameters under the wavelength, PCA showed that plantlets of O. majorana under monochromatic red light had higher arbutin content, and plantlets under white light had higher LDW, SDW and ShDW. The plants showed higher levels of carotenoids and total chlorophyll under green light (Fig. 9A). Regarding light intensity, it is possible to conclude that O. majorana under higher intensity (130 µmol m− 2 s− 1) had higher arbutin content. Plantlets grown under 94 µmol m− 2 s− 1 light intensity showed higher LDW, SDW, ShDW, and SL, while lower light intensity (26 µmol m− 2 s− 1) accumulated a higher content of photosynthetic pigments (chlorophyll a, chlorophyll b and total chlorophyll) (Fig. 9B). The PCA results corroborate and complement the previous analyses and interpretations.
{kind=link}