Examination of molecular interactions
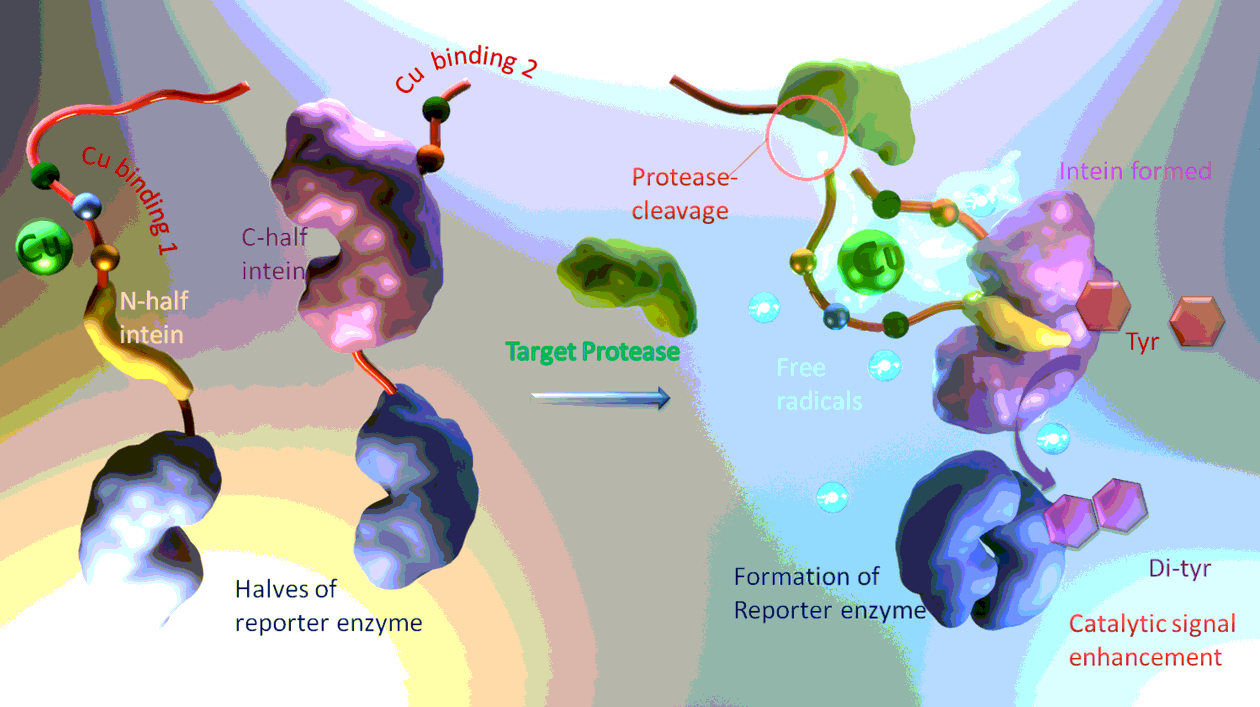
The proposal involves several interactions between proteins, peptides and metal cofactors alike. The design arranges these interactions in an ordered fashion to convert protease activity into enzymatic signal output. However, to realize this design, as depicted in Scheme 1, every interaction evolved should be able to first proceed smoothly by its own, and then their synergic effect then have to be examined to see whether or not all the molecular partners can interact in an orchestrated manner. The interactions in this design are of two types: first, non-covalent molecular recognition between peptide/metal ion and proteins bring the potential molecular partners together, in a loosely associated non-covalent complex, and then covalent bonding and cleavage take place to form the reporter enzyme, as depicted in Scheme 1.
The non-covalent interactions are first studied using isothermal titration calorimetry (ITC) which measures the heat given out upon formation of a non-covalent complex. First, the interaction between the target protease and its substrate peptide is studied. As just being mentioned above, the proteolytic cleavage is actually two steps in tandem: the lock-and-key like molecular recognition of the substrate peptide sequence brings this short peptide to the substrate-binding pocket of the target protease, and then comes the cleavage of bond under proper chemical conditions. Without cofactors, the target protease can only recognize the substrate, forming non-covalent complex. So the formation of this protease-substrate complex is first studied thermally, and the results, as shown in Figure S1, represent a saturated binding of typical 1 to 1 molecular ratio. This result confirms that the target protease can recognize its substrate effectively. Then, the Cu ion binding short peptides are examined for their capability of Cu coordination. As shown in Figure S2, since two short peptides can bind with one Cu ion, a 2-to-1 binding ratio can be observed in the binding curve. Lastly, the interaction between the two halves of Intein is examined, in the absence of any “load” that can be covalently pieced together by intein. As shown in Figure S3, the two halves of intein can recognize each other in a 1 to 1 manner.
Next, these non-covalent interactions are studied in the context of the proposed sensing design: the several functional peptides and proteins are linked together to form the sensing apparatus, can functioning of this apparatus is studied to see if there is mutual interference and hindrance between the various functional modules. First, the two halves of the intein are each loaded with one half of the reporter enzyme, and each has also been elongated with a Cu-binding peptide, but the substrate peptide of the target protease is absent. Then in the absence of cofactors and buffer ions critical for intein-induced peptide bonding, the interaction of the two halves of intein is again examined. As shown in Fig. 1a, the intein can still effectively form in the presence of the Cu binding sequences, although the apparent affinity is somewhat lower. Then, one half of the intein is pre-incubated with Cu ion, and the two reporter-enzyme-loaded halves are allowed to interact again. As shown in Fig. 1b, the apparent affinity is even higher than in the case of the unloaded intein. The apparent binding ratio is still 1: 1, since Cu has already been pre-loaded to one half of the to-be-formed intein. The enhanced binding might be due to the Cu coordination’s facilitating effect. Then, by elongating one of the two Cu-binding peptides, appending to the two halves of intein, with the substrate sequence of the target protease, the protease-substrate interaction, similar to Figure S1, is studied. Shown in Fig. 1c, the target protease can still effectively recognize its substrate peptide, which is now a part of the sensing molecular machine. Meanwhile, the addition of the substrate sequence can also block the intein formation, as shown in Fig. 1d.
Then the covalent interaction, or the formation of the reporter enzyme after the cleavage of the substrate “cap” by the target protease, can be examined by the reporter enzyme’s catalytic activity. First, in the presence of mixed and surplus amount of the target protease, the two halves of loaded intein of varying concentrations are employed to show the activity of the reporter enzyme. Specifically, the two halves of intein is not proteins of equal size, but one of them is just a peptide strand of 16 amino acids [17]. This “short” half, with its C-terminal being elongated with one half of the reporter enzymes, and its N-terminal modified with one of the Cu binding sequence tandem the substrate sequence of the target protease, is added to a mixture of the other half intein (loaded with the other half of the reporter enzyme, and modified by the other Cu binding sequence) and the target protease, both are of mixed concentration. The addition of the short half of intein initiates the protease cleavage of the substrate “cap”, freeing up the short half to insert into its specific pocket on the larger half of intein. The formation of intein then triggers the formation of the reporter enzyme, which can oxidize tyrosine, and this oxidative process can be captured electrochemically as current-voltage peak signals. As shown in Fig. 2a, the detected presence of the catalytic product, or cross-linked tyrosine, the reduction of which is recorded as the electrochemical signal, is proportional to the concentration of the short half of intein added to the mixture, inferring that the above examined split intein-based biosensing molecular machine can work as designed.
Another interesting aspect of the sensing process is that the reporter enzyme alone is not able to oxidize the signal molecule tyrosine. The bi-functional Cu ion is needed: As a control, the split intein-based sensing molecular machine, without pre-complexed Cu ion, is used in detecting 10 nM target protease, and after incubation of the short half intein to the mixture of the larger half intein and the target protease, Cu ion is then added to the reaction system for tyrosine production. Then, the time course of the whole sensing process is recorded as Figure S4a, compared with similar time course using Cu ion complexed split intein, shown as Figure S4b. It can be found that without first putting Cu ion on one half of the intein, the two halves can still recognize each other after enzymatic cleavage of the substrate peptide, indicated by the existence of tyrosine signal. For if the split intein, without the help of Cu ion and its two binding peptides, cannot come together by themselves, then the reporter enzyme would not form, and the signal readout would not be detected at all. However, the time course in the absence of pre-existed Cu ion on the split intein, is evidently more sluggish than that shown in Figure S4b, indicating that without the help of Cu and its binding peptides, the split intein may take longer to piece together. The second function of Cu ion is facilitating the catalytic oxidation of tyrosine. This can be examined electrochemically, as shown in Fig. 2b. As the potential is cycled, the oxidative and reductive peaks of Cu ion can be recorded. The oxidation of tyrosine catalyzed by the reporter enzyme needs peroxide species or reactive oxygen, as the oxidant, while this oxidant can be generated by the coordinated Cu ion, via the change of valence of that ion, and this change can be recorded as Fig. 2b.
Analytical performance
Under these optimized conditions, the quantitative detection of the target protease in complicated mixture, i. e. in serum diluted samples can be achieved (Fig. 3a). The tyrosine oxidative signal is proportional to target concentration. And the relation between peak current signal level and protease concentration is shown in Figure s3b. The specificity of detection is also satisfactory (Fig. 3b), with respect to several major component proteins of human serum.
Detection of clinical samples
Serum samples from patients of type II diabetes, as well as from healthy volunteers, are collected and assayed to examine possible existence of the target protease. The results are summarized in Fig. 4, and the samples are divided into three groups: the control group represents the healthy volunteer, while the patients with jobs mainly working in an office in the last decades compose the “in door” group, which may be subject to less exposure to air pollutants on the streets. In contrast, the patients holding outdoor jobs are grouped into the “outdoor” group, which might have endured more air pollution than the indoor group. Overall, the target protease is not abundant in serum. This is evident for the healthy volunteers whose protease level is almost below the limit of detection of the proposed method. Diabetes statistically significantly elevates the serum level of the target protease, although the absolute concentration is still not significant. The out-door group indeed displays some elevation of the target protease, if compared with the in-door group, although the difference is not too statistically significant.
{kind=link}