In breast cancer patients with T2D, use of metformin during neo-adjuvant chemotherapy has inhibitory effects on tumor progression (48). Metformin also reduces the tumor growth in non-diabetic patients with obesity with the mildest side effects (26, 49). Epidemiological studies have shown that diabetic women treated with metformin, but no other anti-diabetic drugs had lower breast cancer incidence compared to the non-diabetic women (22). This association was specifically observed for ER+, PR + and HER2- (luminal A) breast cancer subtypes (50, 51). These findings suggest a direct link between T2D and subtype-specific breast cancer that might benefit from metformin treatment. In the present study, most of the participants were non-diabetic overweight/obese ER- breast cancer survivors (triple negative and non-luminal HER2+, Table S1). As there was no significant difference in the tumor subtype history in each treatment arm, this study is not able to differentiate the preventive effect of metformin specific to a subtype of breast cancer.
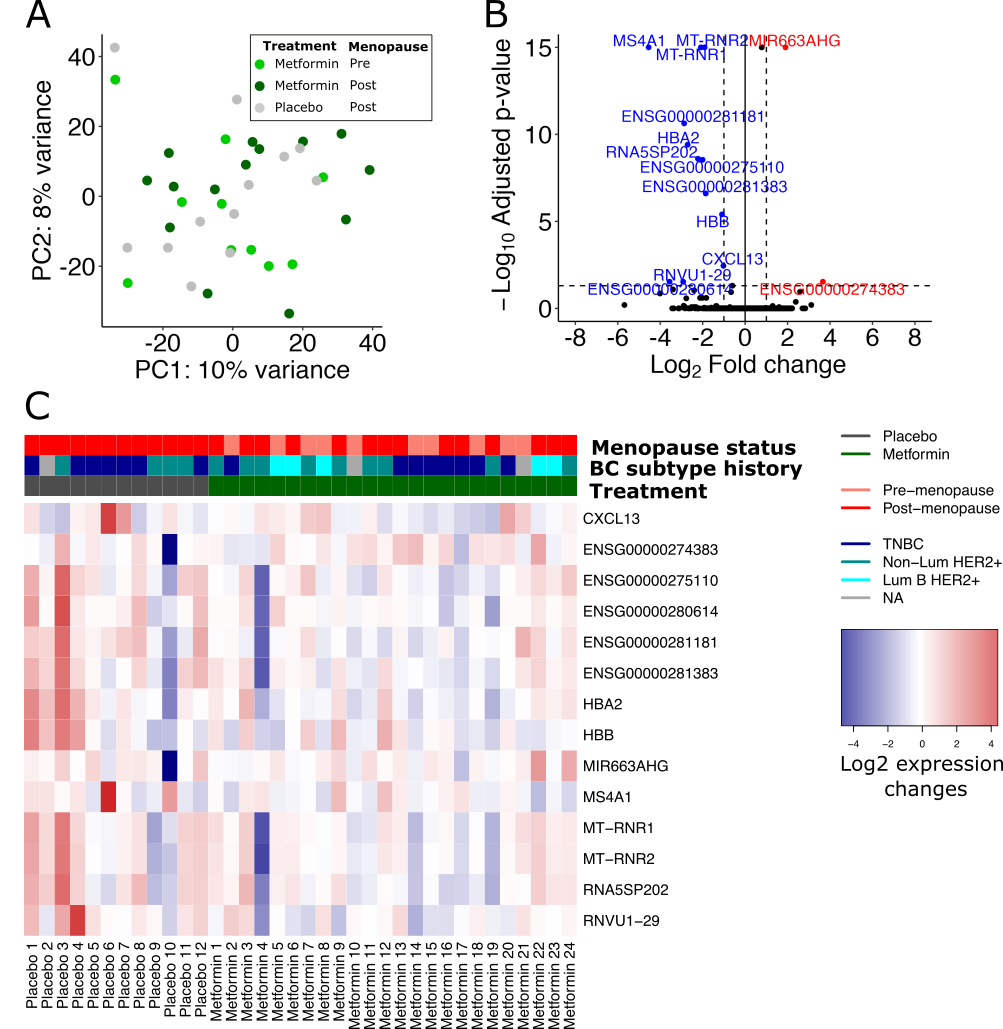
As far as we know, our study is the first to investigate whether metformin may have a preventive effect on the initiation of new tumor lesions in breast tissue by RNA sequencing analysis. Here, we employed global gene expression analyses of the breast tissue biopsies from placebo- and metformin-treated participants at baseline and post-treatment. By comparing metformin vs. placebo-treated breast tissue transcriptomic profiles, we identified several down-regulated genes including protein-encoding genes MS4A1, HBA2, and two mitochondrial genes MT-RNR1/2. Changes in MS4A1 expression in breast tumors have been related to lower lipid metabolism and better survival (40). MS4A1 also encodes a B cell and a T cell subset surface marker (CD20) and is closely related to the immune-active tumor microenvironment and immune cell activation-related pathways in breast cancer (40). Hemoglobin genes including hemoglobin subunit β (HBB) and HBA2 are biomarkers of inflammatory breast cancer (41). A transcriptomics study on the blood cells of the healthy individuals showed that short-term administration of metformin decreased the expression of HBA and HBA2 (52), a metformin-associated effect that we also observed in our study on whole gene expression profiles of breast tissue of the breast cancer survivors. The transcripts encoded by MT-RNR1/2 genes have been associated with hepatocellular carcinoma and triple negative breast cancer progression (44). The two MT-RNR1 and MT-RNR2 not only encode ribosomal RNA 12S and 16S, respectively, but are also transcribed to small proteins that are crucial for metabolic homeostasis. MT-RNR1 encodes a peptide called mitochondrial open reading frame of the 12S rRNA-c (MOTS-C) that is known to regulate insulin sensitivity through AMPK signaling pathway (53). MT-RNR2 also encodes a conserved cyto-protective polypeptide called humanin that promote tumor progression and metastasis by its anti-apoptotic effects (44).
Whether overweight and obesity increase the risk of breast cancer in premenopausal women is controversial (54). Due the difference in the expression profiles and possibly a different molecular mechanism of metformin’s actions on pre- and postmenopausal breast tissue (39, 55), we are not able to extend the previously mentioned findings of the studies to the premenopausal women. In the present clinical trial, all the premenopausal breast cancer survivors were randomized to metformin-treatment arm, preventing us from studying the potential molecular effects of metformin in the premenopausal overweight/obese breast cancer survivors.
In the differential analysis where changes in expression exclusively from postmenopausal breast tissue samples were examined, we found two additional down-regulated oncogenes, EGFL6 and FDCSP. EGFL6 is highly expressed in various malignant tumors compared to normal tissues (56, 57). EGFL6 is associated with cancer cell proliferation, migration, invasion and angiogenesis (58), and promotes epithelial to mesenchymal transition of the cancer stem cells (59), a mechanism targeted by metformin in breast tumors (60, 61). FDCSP is highly expressed in epithelial ovarian, breast, endometrial and prostate cancer types (62) and contributes in the cancer cell invasion and metastasis through Akt signaling and down-regulation of E-cadherin (62). Despite the limited knowledge of how these genes might be involved in regulation of cancer initiation and tumorigenesis, these transcriptomics data suggests that metformin might induce expression profile in normal breast tissue that precludes development of new tumors.
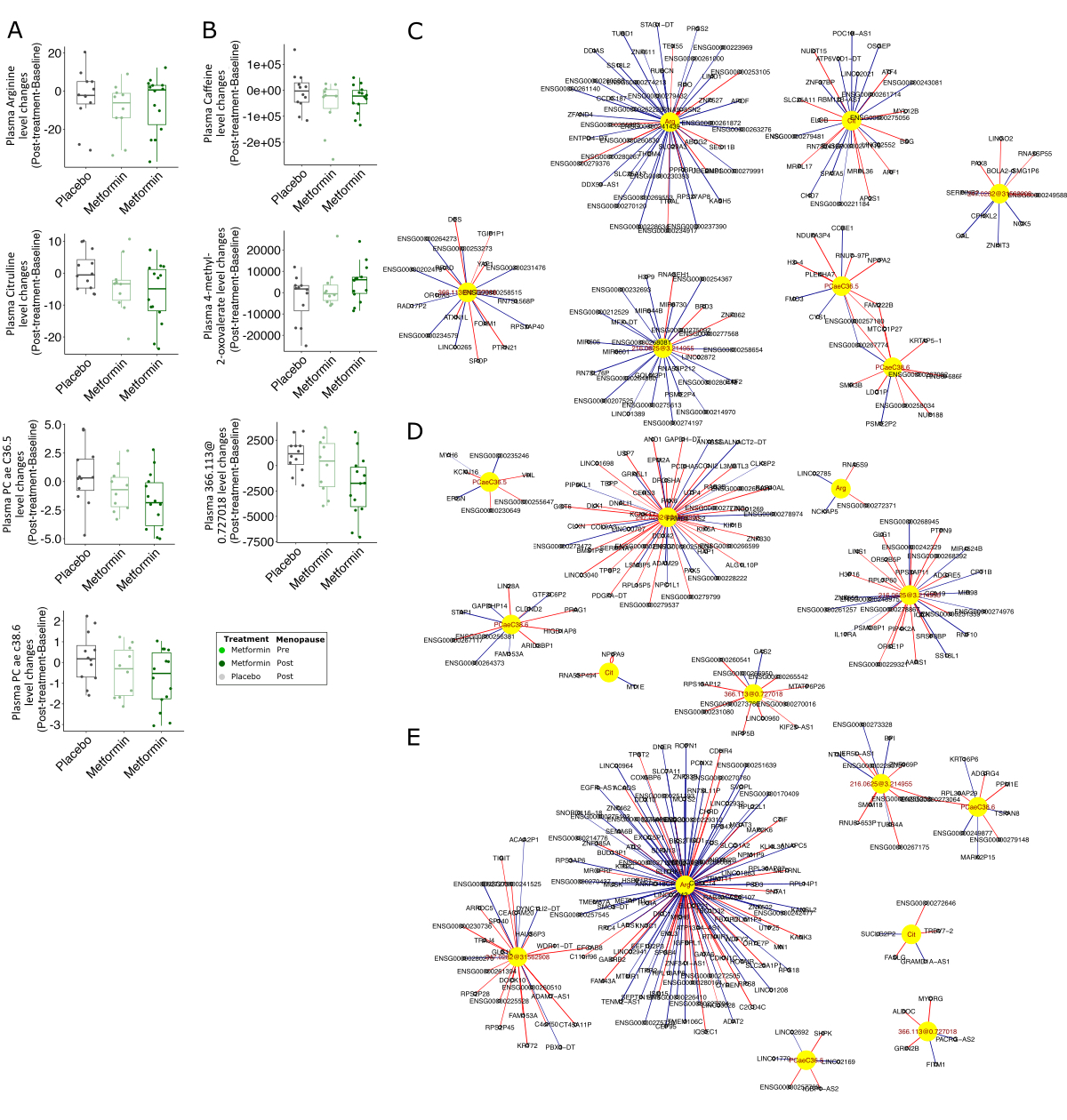
The metabolic reprogramming in cancer cell is essential to provide the energy need for rapid growth and proliferation of cancer cells (63). Metabolic changes may also induce dormant breast cancer cells to change their quiescent state (64). One of the possible mechanisms of metformin action at the cellular level is disrupting mitochondrial function, thereby resulting in alterations of citric acid cycle (65), oxidative phosphorylation as well as ATP production (66). In this study, we employed metabolomics approaches to explore the potential associations between metabolites and breast tissue gene expression patterns in metformin-treated breast cancer survivors (31). Using targeted metabolomics on the plasma samples, we observed metformin-associated changes in acylcarnitines, amino acids, lysophosphatidylcholines and several phosphatidylcholines, some of which were also reported to be similarly changed following metformin administration in the previous pooled RFH + MetBreCS study (31). Carnitine and acylcarnitines are essential compounds for the oxidative metabolism of fatty acids in the mitochondria. Abnormal acylcarnitine metabolism has been associated to insulin resistance in T2D, obesity and breast cancer (67, 68). In contrast to the pooled RFH + MetBreCS study (31), we found a slight metformin-associated reduction in malonylcarnitine/hydroxybutyrylcarnitine, and an increase in acylcarnitines dodecanoylcarnitine and tetradecadienylcarnitine, which are all known to be increased in plasma and urine from pre-diabetic and T2D individuals (69, 70). We also found a decreased trend in the plasma levels of long-chain unsaturated phosphatidylcholine species, two of which (PC ae C36:5 and PC ae C38:6) were also significantly decreased with the metformin treatment in the pooled RFH + MetBreCS study (31). While PC ae C38:6 has been reported to be strongly lowered by metformin treatment in patients with T2D (71), metformin leads to increase in the levels of other phosphatidylcholines (72, 73). Metformin also reduces cellular levels of several lysophosphatidylcholines, such as lysoPC a C16:0, lysoPC a C18:0, lysoPC a C18:1 and lysoPC a C18:2 (74, 75). However, in our study, two lysophosphatidylcholines (lysoPC a C18:1 and lysoPC a C20:4) were increased in the plasma of metformin-treated breast cancer survivors. Although the tissue-specific effects of metformin on phospholipid metabolism is not well studied, decreased phosphatidylcholine production in breast cancer cells is attributed to reduced activity of phosphatidylcholine-synthesizing enzymes choline kinase and CTP:phosphocholine cytidylyl transferase, and increase activity of phospholipid-specific phospholipase C (73). Here, we did not observe any change in these gene transcripts or significant correlations with plasma changes of phosphatitylcholines in the metformin group. However, it should be noted that plasma measurements do not necessarily correspond to breast cell microenvironment. Whether the metformin-associated changes in the plasma levels of phosphatidylcholines correlate with the actual concentrations within the breast tissue remains to be examined.
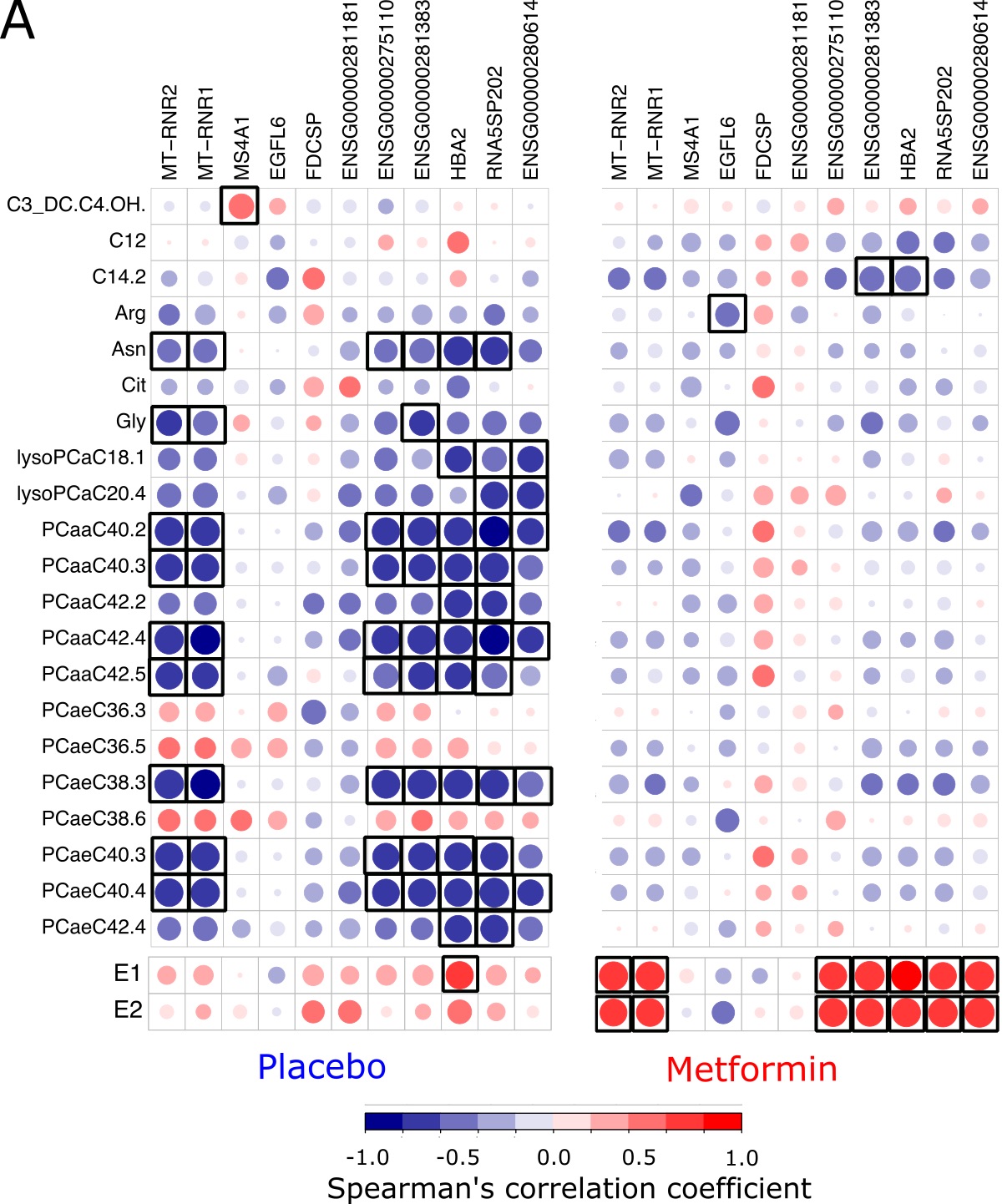
Arginine, which is synthesized from citrulline regulates metabolic processes such as synthesis of polyamines and nitric oxide (76), the latter of which modulates different cancer-related events (77). Although the arginine-derived polyamines spermine and spermidine were excluded from our analyses, there is evidence that up-regulation of polyamines synthesis due to increased levels of arginine in the tumor microenvironment leads to DNA methylation and thus repression of genes encoding pro-inflammatory cytokines in the immune cells (78, 79). Plasma arginine level is also associated with enhanced innate and adaptive immune responses in the tumors (80, 81). Dietary supplementation with L-arginine alone or in an adjuvant setting in patients with breast cancer significantly enhances host defenses by natural killer cells and activated lymphocyte cell cytotoxicity (82, 83). On the other hand, arginine deprivation leads to decreased cell growth and proliferation, and cell death in many tumor types (45, 84). Our functional analysis on the integrated gene expression profile and plasma arginine and citrulline levels led us to identify the associated signaling pathway in breast tissue. Compared to the placebo-treated group, the metformin-associated decrease in plasma arginine and citrulline revealed enrichment of reduced immune responses, mitochondrial oxidative phosphorylation and endothelial cell proliferation and migration pathways.
In this study, we also observed a trend of decreased plasma caffeine levels in participants receiving metformin, in line with the previous observation in the metformin-treated population of the pooled study (31). Although caffein consumption details were not collected from in the participants, a higher activity of hepatic CYP1A2 would explain these changes (31). Such a change in CYP1A2 activity might be an indirect effect of metformin on reducing pro-inflammatory cytokines (85) and/or an increase in CYP1A2 expression and activity (86). Despite the importance of CYP1A2 in the pathogenicity of breast cancer (87), CYP1A2 expression is very low in breast tissue (less than 10 read counts through all samples in our data set) and therefore it was excluded from our analyses.
The levels of several sex steroids are related to breast cancer risk and prognosis in postmenopausal women (88). Elevated serum E2 is linked to increased risk of breast cancer particularly in obese postmenopausal women (11). In premenopausal women, circulating E2, mainly synthesized in the ovaries, may take part in the development and growth of breast cancer cells. After menopause, estrogens are mainly generated by extragonadal organs through conversion of androstenedione and testosterone to E1 and E2, which is catalyzed by the aromatase (15). Deregulation of serum concentrations of E1 and E2, mainly through the higher activity of aromatase enzyme in the visceral adipose tissue combined with decreased liver production of SHBG, has been suggested as the main mechanism that connects postmenopausal obesity and T2D and breast-cancer risk (16, 89). In line with previous studies, we show that administration of metformin to postmenopausal breast cancer survivors reduces the serum levels of E2 (90–92). Although the mechanisms by which metformin reduces the systemic levels of estrogen is not well understood, the increased expression of LKB1 and activity of AMPK, cytoplasmic sequestering of the CREB co-activator and consequently down-regulation of aromatase expression in the adipose tissue might explain the reduced levels of estrogen in the postmenopausal women (93, 94). There are also evidence suggesting that the aromatase gene (CYP19A1) expression is targeted by metformin through the PII/PI.3 promoter (95). In our RNA seq data, we were not able to detect any changes in CYP19A1 expression in breast tissue from metformin-treated women. However, we observed negative correlations between the CYP19A1 expression and changes in serum androstenedione and testosterone in the placebo- and metformin-treated arms, respectively, but no correlations between CYP19A1 and changes in serum E1 or E2 (Fig. S4A). Nevertheless, the expression of CYP11A1 and CYP1B1 were negatively correlated with the changes in serum E1 and E2 only in the metformin-treated postmenopausal group. The negative correlations between CYP11A1 and CYP1B1 expression and the sex hormones E1 and E2 might suggest a feedback mechanism to reduce the biosynthesis of estrogens and further metabolism to DNA damaging quinones. Although we did not find any changes in the mRNA expression of these enzymes in the breast tissue of the metformin-treated group, it has been reported that metformin is able to directly reduce the protein levels of CYP11A1 in the ovary granulosa cells (96) and CYP1B1 in breast cancer cells (97).
The present study has several limitations; first, the transcriptomics analysis is performed on the global gene expression, and we may not conclude on cell-specific metformin effects. Moreover, the analyses of sex hormones and metabolites are limited to the systemic levels and do not provide a tissue-specific perspective. However, the integration of the systemic E2 and the altered metabolites with the corresponding gene expression profile in the breast tissue provides an overall view in the treatment effect of metformin. Furthermore, the functional analyses were performed based on correlation analyses with no adjustment for any confounding factors such as baseline values and BMI and cannot prove causality but only show associations. Lastly, the small sample size in MetBreCS trial limits the statistical power in our analysis.
{kind=link}
{kind=link}
{kind=link}
{kind=link}