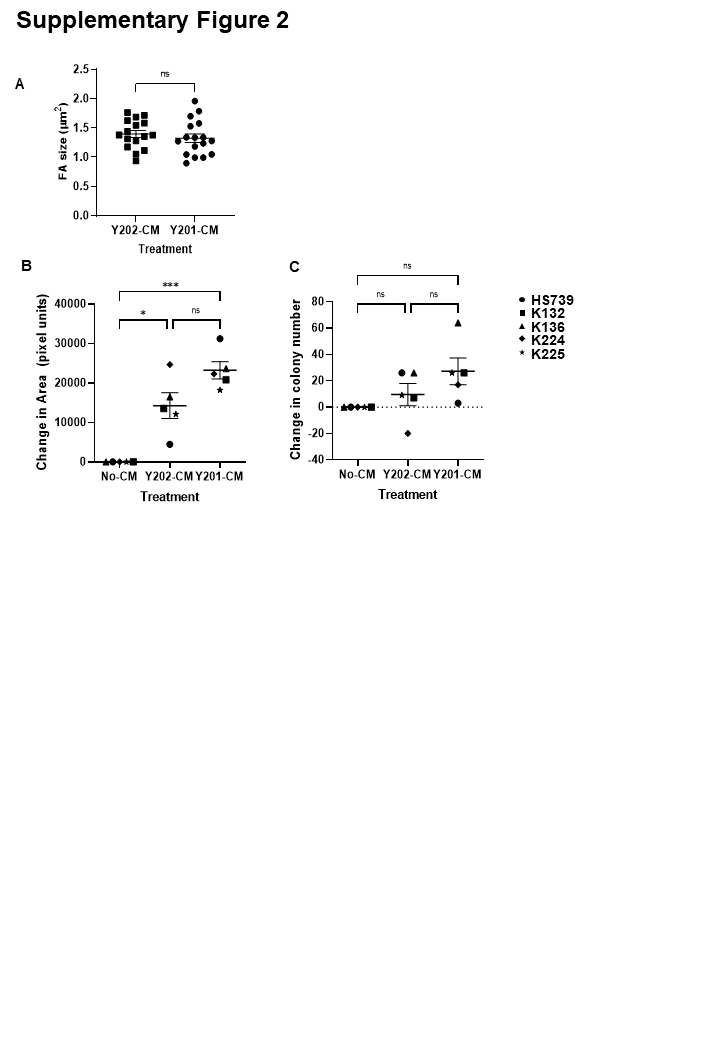
Our findings provide further evidence of a correlation between BMSC morphology and functionality, supporting previous evidence that morphologically distinct stromal subsets are likely to reflect functional heterogeneity, and observations that cells with different morphologies have altered inflammatory or differentiation characteristics (19, 37–40) We exploited a label-free ptychographic technique to track the morphology and motility of cells over time (41). This could prove useful in the real-time discrimination of primary cell population phenotypes without the need for fluorescence-based or other end-point labelling methods. Using our simplified model of BMSC heterogeneity we showed that large, flat, inflammatory BMSCs were less motile than stem cell-like, spindle-shaped cells. In addition, cells matching these morphological parameters were reproducibly observed in primary cultures, suggesting that image-based morphometric analysis could be employed as a predictive measure of cell function, with previous evidence suggesting faster migrating BMSCs are indeed more likely to be multi-potent (42). Furthermore, we demonstrated that the morphometric features of the atypical flattened BMSCs (Y202) were plastic and could be modified by exposure to factors secreted by more typical, spindle-shaped Y201 BMSCs. BMSC phenotypic plasticity may be, to some extent, determined by the secreted factors of the cell population as a whole, with ECM components being important determinants of cell behaviour.
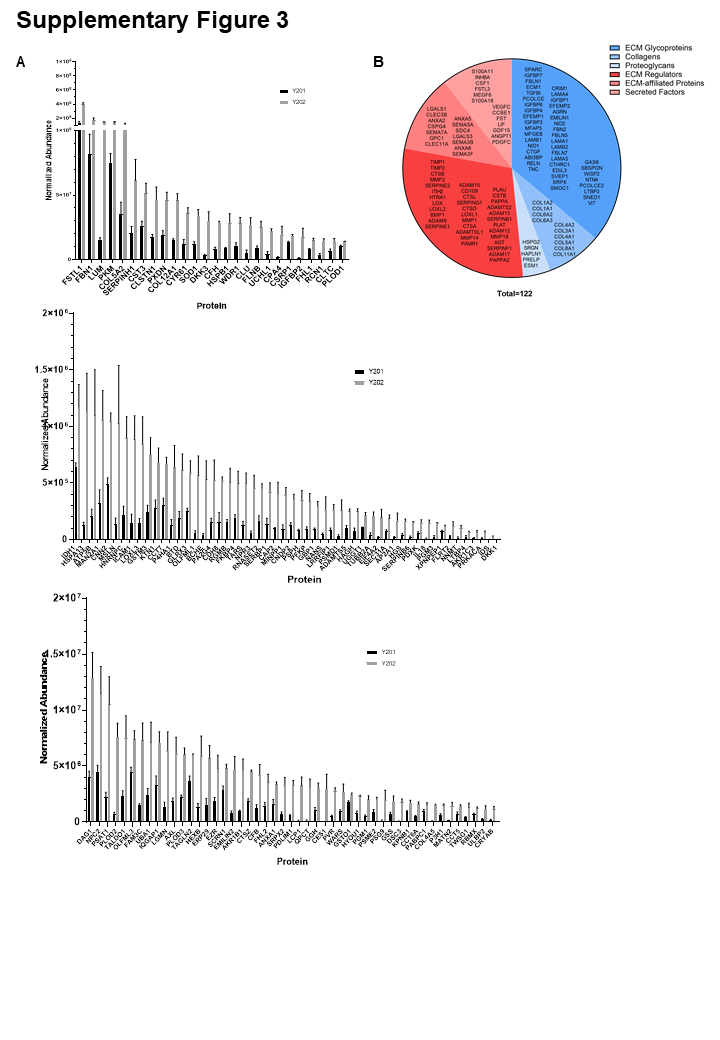
The BMSC secretome is linked to cellular functionality, which is important both for the understanding of disease and potential uses of these cells in therapies (43). We found that the secretome of multipotent Y201 BMSCs was strongly enriched for proteins involved in the production and modification of the ECM, as well as TGF-beta and Notch signalling pathways both of which are implicated in controlling BMSC differentiation (44, 45). Subsequent assessment of the ECM produced by these BMSC lines identified a thicker and more complex matrix produced by Y201 cells, while Y202-derived matrices were relatively thin. The ECM has a prominent role in driving migration, and as such the increased production and secretion of matrix proteins captured in conditioned media could contribute to the phenotypic switch we saw in Y202 cells. Lumican, which was secreted at higher levels by Y202 cells, has previously been shown to inhibit the migration of MSCs, as well as regulation of immune responses in other cell types, potentially correlating with the slow moving immune-based role of Y202 (46).
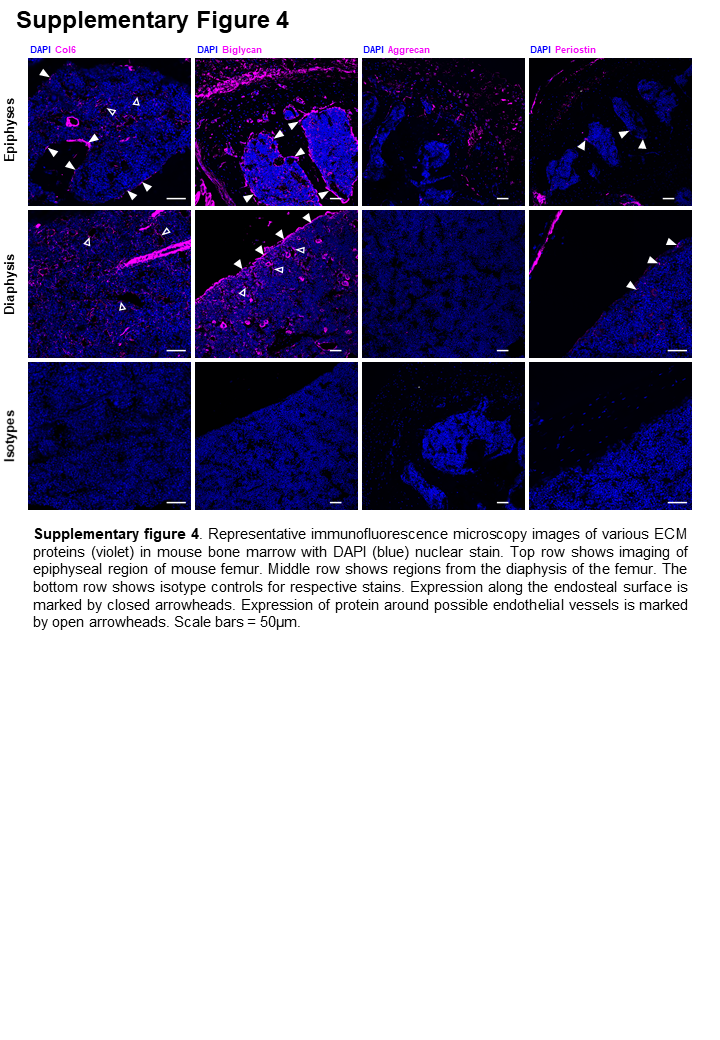
Similarly, aggrecan and periostin which were more abundantly secreted in Y201 BMSCs compared to Y202 BMSCs, may act as candidate differentiators of cell phenotype. It is also possible that those proteins that did not differ significantly between the two BMSC lines represent a ‘core’ matrix common to all BMSC subtypes. The ECM is undoubtedly important for cellular function, mediating biochemical and mechanical signals to cells. Molecular patterning of a niche environment has previously been shown to regulate macrophages between a pro-healing and inflammatory phenotype (47). This is likely to be of high importance for stem cells in a structurally diverse tissues such as bone marrow, where the role of ECM in maintaining hematopoietic stem cells in their niche has been increasingly characterised (48, 49). Our identification of aggrecan and periostin underlying CD271+ cells in human bone marrow provides promising evidence that an in vitro matrix produced by cells isolated from a complex tissue may, at least in part, recapitulate the in vivo ECM composition, and may indeed contribute to a specialised stromal niche. Our findings are supported by previous evidence for CD271+ BMSC niches, with CD271+CD56+ cells found exclusively on trabecular bone surfaces, representative of an endosteal BMSC niche (50, 51). CD271+ BMSCs are also 65-fold increased in BMSCs isolated from trabeculae versus bone marrow aspirates, again highlighting a more endosteal niche for this population (52). The same pattern of CFU-F capacity and in situ localisation is seen when combined with another prospective potency marker, melanoma cell adhesion molecule (MCAM/CD146), as CD271+CD146−/low populations were found as bone lining cells, whereas CD271+CD146+ were located perivascularly (53). We hypothesise that differentiation-competent cells pattern their local environment with a phenotype-supportive matrix that is physically and biochemically suited to cell function, with our findings complementing other studies showing that matrix of young MSCs has been shown to restore proliferation and differentiation to older MSCs which has important implications for downstream therapeutic development (54–56).
Periostin has previously been linked with controlling the regenerative potential of periosteal skeletal stem cells, as well as supporting haematopoietic stem cells in the foetal liver niche and regulating their quiescence (57, 58). The observation of rare periostin in bone marrow has not been previously reported in large-scale analyses of protein distribution across whole long-bones, however BMSC-derived periostin has also been shown in mouse to have functional effects in leukaemia, suggesting it is present in marrow (58–60). Further, periostin knockdown in human BMSCs results in inhibition of osteogenic differentiation of these cells, indicating its importance for a differentiation-competent, stem cell phenotype.(61) The observation of periostin and aggrecan expression in trabecular bone regions in mouse and human tissue sections might also indicate conservation across species for these proteins in a stromal niche for bone lining cells. Follow up work to isolate CD271 + aggrecan and periostin-expressing primary BMSCs is necessary to determine if these possible biomarkers of potency are consistent and selective.
We also demonstrated that the ECM substrate produced by Y201 BMSCs may be partly responsible for its enhanced migratory phenotype; exposure of Y202 BMSCs to ECM derived from Y201 BMSCs resulted in increased migration, speed and distance in a FAK-dependent manner. The ability of the Y201 matrix to instruct the immotile Y202 to migrate highlights the importance of the secreted ECM in phenotypic plasticity and could correlate with previous observations of rejuvenation of cells by exposure to specific ECMs.(55)
{kind=link}
{kind=link}
{kind=link}
{kind=link}