3.1. Preliminary phytochemical screening for B. monnieri and C. asiatica extract extracts
3.1.1. In vitro antioxidant activity
(i) Total antioxidant activity
The total antioxidant capacity (TAC) assay was done based on the Phosphomolybdenum method [18], here the sample reduces Phosphate-Mo (VI) to Phosphate Mo (V) and forms a bluish-green colored phosphate/Mo (V) complex. The present study observed that BM aqueous extract showed higher TAC activity, 56.56 ± 1.55, than CA aqueous extract, i.e., 45.68 ± 0.21 at 800 µg/ml concentration (Fig. 1A). The TAC was quantified as ascorbic acid equivalents. The TAC activity is directly proportional to the concentration of B. monnieri and C. asiatica extract.
(ii) 2,2-diphenyl-1-picrylhidrazyl (DPPH) free radical scavenging activity
B. monnieri and C. asiatica aqueous extracts showed significant DPPH free radical scavenging activity. Here the DPPH nitrogen radical accepts hydrogen from an antioxidant and leads to the disappearance of the DPPH nitrogen radical in the test sample [45]. The change in color from purple to yellow shows the disappearance of the DPPH radical at the wavelength of 517 nm [19]. B. monnieri aqueous extract showed the highest free radical scavenging activity at 400 µg/ml concentration (Fig. 1B), i.e., 80.05 ± 0.05, which remains constant. On the other hand, C. asiatica extract showed less free radical scavenging activity when compared to B. monnieri aqueous extract, i.e., 54.90 ± 2.34 at 800 µg/ml.
(iii) Reducing power assay:
This assay measures the sample’s reductive ability, indicating its potential antioxidant activity. The reducing power of B. monnieri and C. asiatica aqueous extract increased with the sample concentration. Among the B. monnieri and C. asiatica aqueous extracts, B. monnieri aqueous extract showed the highest reducing power at 800 µg/ml concentration, i.e., 55.35 ± 0.39, compared to the C. asiatica, i.e., 21.65 ± 0.52. Ascorbic acid was taken as a standard for calculating reducing power. Figure 1C represents the reducing power of the B. monnieri and C. asiatica aqueous extract.
3.1.2. Quantitative analysis of secondary metabolites
(i) Phenolics
According to Kahkonen et al. [46], phenolic compounds directly contribute to the antioxidant property of plant extract. Therefore, we tested the phenolic content of both B. monnieri and C. asiatica aqueous extracts in the present study. Our results showed that as the sample concentration increases in µg/ml, the total Phenolic content (µg/ml) also increases. Figure 2A represents the total Phenolic content in the B. monnieri and C. asiatica aqueous extract. The total Phenolic content of B. monnieri and C. asiatica aqueous extract at 800 µg/ml is 107.18 ± 1.28 (P < 0.0001) and 46.03 ± 0.41 (P < 0.0001), respectively, and 1000 µg/ml is 130.64 ± 0.16 (P < 0.0001) and 55.39 ± 1.31 (P < 0.0001) respectively.
(ii) Flavonoids
Flavonoid is a class of secondary metabolites having a polyphenolic structure. It is associated with health-promoting effects such as antioxidant, anti-inflammatory, anti-mutagenic, and anti-cancer [47]. B. monnieri and C. asiatica aqueous extracts showed high flavonoid content in µg/ml. For example, the flavonoid content of B. monnieri and C. asiatica aqueous extract at 800 µg/ml were 4.44 ± 0.19 (P < 0.0001) and 7.55 ± 0.22 (P < 0.0001), respectively. At 1000 µg/ml concentration, the flavonoid content was 5.52 ± 0.09 (P < 0.0001) and 9.09 ± 0.03 (P < 0.0001), respectively. Figure 2B represents the total flavonoid content in the aqueous extract. Here, quercetin was taken as a reference standard.
3.2. Metabolite identification for B. monnieri and C. asiatica extracts
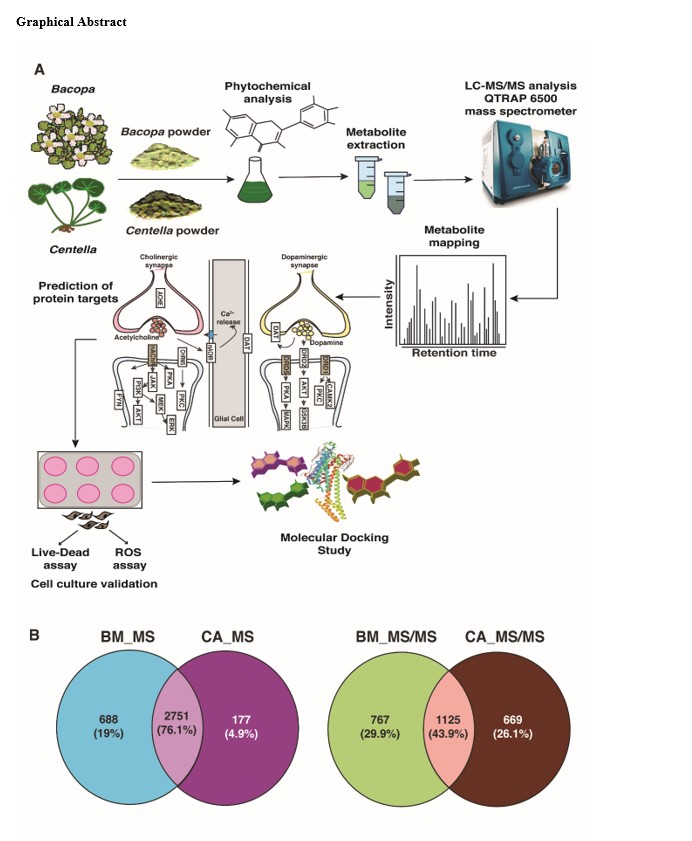
The small molecule metabolites are the players at the cellular level, conferring neuroprotection. We used the untargeted metabolomics approach to unravel and compare the molecular cues using PlantCyc and KEGG databases. The total ion chromatogram of B. monnieri and C. asiatica from positive and negative modes is given in Supplementary Fig. 1. Metabolite identification was performed at the precursor level for B. monnieri and C. asiatica. Analysis of B. monnieri revealed 2,614 and 2,849 metabolites in positive and negative modes, summing to 3,438 non-redundant metabolites. In C. asiatica, we identified 2,826 and 778 metabolites in positive and negative modes, respectively, representing 2,927 non-redundant metabolites. Finally, we compared the metabolite profiles across B. monnieri and C. asiatica, wherein 2,751 (76.1%) metabolites were shared. The mapped metabolite features with respective intensities and polarities from B. monnieri and C. asiatica are given in Supplementary Tables 1–4.
We also identified metabolites at MS/MS level using our in-house tool, MS2Compound [26]. As a result, we identified 1892 and 1794 metabolites from both the polarities for B. monnieri and C. asiatica, respectively. Among these, 767 and 669 metabolites were unique for B. monnieri and C. asiatica, respectively, with 1125 metabolites common between them. These metabolite names at MS/MS level are given in Supplementary Table 10.
The mapping of metabolites is purely based on the database used. Despite using two different databases, a large amount of data from B. monnieri and C. asiatica remained unassigned, as provided in Supplementary Tables 5–8. For example, at the MS1 level, 5741 and 4161 m/z remained unassigned for B. monnieri and C. asiatica, respectively.
3.3. Identification of previously reported metabolites from B. monnieri and C. asiatica extracts
We looked for the previously reported metabolites of B. monnieri and C. asiatica at precursor and fragment-level from the literature. In the case of B. monnieri, we could map phytochemicals such as sitosterol, betulinic acid, and L-aspartate at precursor and fragment levels and D-mannitol, stigmasterol, L-alanine/α-alanine at MS1 level as reported by Jeyasri et al. [48]. For C. asiatica, we could map Asiatic acid at the precursor level as previously reported [49], alpha-humulene, β-Caryophyllene, and campesterol at the precursor and chavicol [50] at both precursor as well as fragment level (Table 1). The spectra of these metabolites are given in Fig. 3A-B. Identifying previously reported metabolites gives us confidence and validates our data and pipeline used for experiments.
Table 1
List of some previously reported metabolites represented as signature metabolites for Bacopa monnieri and Centella asiatica
|
Plant extract
|
Metabolite name
|
Molecular formula
|
Monoisotopic mass
|
MS1/ MS2/
MS1 & MS2 (both)
|
|---|
|
BM
|
Sitosterol
|
C29H50O
|
414.38
|
MS1 & MS2
|
|
Betulinic acid
|
C30H48O3
|
455.35
|
MS1 & MS2
|
|
L-aspartate
|
C4H5NO4
|
134.04
|
MS1 & MS2
|
|
D-mannitol
|
C6H14O6
|
182.07
|
MS1
|
|
Stigmasterol
|
C29H48O
|
412.37
|
MS1
|
|
L-alanine
|
C3H7NO2
|
90.05
|
MS1
|
|
CA
|
Asiatic acid
|
C30H48O5
|
488.35
|
MS1
|
|
Alpha-humulene
|
C15H24
|
204.18
|
MS1
|
|
β-Caryophyllene
|
C15H24
|
203.17
|
MS1
|
|
Campesterol
|
C28H48O
|
400.37
|
MS1
|
|
Chavicol
|
C9H10O
|
134.07
|
MS1 & MS2
|
3.4. Metabolite chemical classification and pathway analysis
The metabolites identified from the B. monnieri and C. asiatica extracts were enriched for their chemical classes, and 94.8% of the classes were similar between B. monnieri and C. asiatica (Fig. 4A). As represented in Fig. 4A, the metabolites mainly comprised alkaloids, flavins and flavonoids, benzenoids, nucleic and organic acids, TCA cycle, phenols, quinones, and carbohydrates. The lipids identified belonged to the classes prenols, sterols, and sphingolipids. Benzoxazines, pyridine carboxylic acids, and glycerophosphates were exclusively present in B. monnieri, while pyranones and ketones were exclusively present in the C. asiatica extract listed in Supplementary Table 11. The enrichment for metabolic pathways resulted in 92.1% similarity. The common and highly enriched pathways between B. monnieri and C. asiatica extracts were salvage pathways for nucleotides and nucleosides, UTP and CTP dephosphorylation, organic nitrogen assimilation, and tetrahydrofolate biosynthesis, to name a few, given in Supplementary Table 12. Enriched pathways related to neuronal functions included choline biosynthesis, tryptophan metabolism, glutamate degradation, MAP kinase cascade, serotonin and melatonin biosynthesis, and dopamine metabolism. The metabolites found in B. monnieri and C. asiatica extracts shared a significant similarity, suggesting they might interact with related proteins. Therefore, based on structural similarity, potential protein targets that bind to the B. monnieri and C. asiatica extract metabolites were retrieved from the BindingDB database.
3.5. Protein targets regulating neuronal functions
B. monnieri and C. asiatica are considered neuroprotectants in the traditional Indian medicinal system; we used the metabolomics data to map their protein targets to evaluate their potential interacting partners. Molecular interaction between the identified metabolites and proteins provided us with valuable information on the probable molecular targets of these nootropics. The BindingDB analysis identified 390 and 370 human protein targets for metabolites corresponding to B. monnieri and C. asiatica, respectively (Supplementary Table 13). 92.4% (365) of proteins were common between both metabolite extracts. Further, these proteins were subjected to functional enrichment analysis to find out the pathways regulated by them. Neuroactive ligand-receptor activation was the most enriched pathway for B. monnieri and C. asiatica protein targets. Other neuronal pathways such as Serotonergic, Cholinergic, and Dopaminergic synapse, Calcium signaling, PI3K-AKT, and MAPK pathway were also enriched (Supplementary Tables 14 and 15). The top 20 enriched pathways for B. monnieri and C. asiatica, respectively, are given in Fig. 4B-C.
3.6 Regulation of neurotransmitter-related signaling pathways
Regulation of neuronal functions
Protein target enrichment enabled us to narrow down important neuronal functions such as calcium signaling, neurotrophin signaling, metabolic pathways, and endocrine resistance. In addition, cellular functions such as cell cycle, autophagy, apoptosis, long-term potentiation, and long-term depression have been implicated in normal physiological functions and various neurodegenerative diseases (Supplementary Tables 14 and 15).
Signaling mechanisms related to neuronal activities
Several signaling mechanisms were enriched that may play an important role in the regulation of neuronal activities, such as AMP-activated protein kinase (AMPK), Vascular endothelial growth factor (VEGF), Ras-related protein 1 (RAP1), Mitogen-activated protein kinase (MAPK), RAS, FoxO, cGMP-PKG, Estrogen and ErbB signaling, and longevity regulation pathways (Supplementary tables 14 and 15).
Neurodegenerative disease-related pathways
According to an increasing number of studies, the development and control of neurodegenerative disorders are significantly influenced by changes in signaling pathways. Therefore, understanding these pathways might result in better disease management. Several enriched protein targets, including CHRM, KRAS, MAP2K1, GSK3B, AKT1, PRKACA, MAOB, ADORA2A, and SLC6A3, among many others in the present study, mainly belong to the most prevalent neurodegenerative diseases, such as Alzheimer’s and Parkinson’s disease, where the cholinergic and dopaminergic neuronal activities are affected. The components of these pathways were identified as potential targets of B. monnieri and C. asiatica metabolites (Fig. 5A-B.). The present results indicate that B. monnieri and C. asiatica may have neuroprotective properties in these diseases as their metabolites may regulate cholinergic and dopaminergic synapses. We also identified many classes of neuroactive receptors and their respective ligands, detailed in Table 2.
Table 2
List of neuroactive ligands and receptors as hits of BM and CA
|
Class-A Rhodopsin-like Amine ligands
|
|---|
|
List of neuroactive ligands and receptors as hits of Bacopa monnieri and Centella asiatica
|
Receptor
|
Proteins identified (Official Gene symbols)
|
|---|
|
Acetylcholine
|
Cholinergic receptors
|
CHRM5
|
|
Epinephrine and norepinephrine
|
Adrenergic receptors
|
ADRA1A
|
|
ADRA2A
|
|
ADRA2B
|
|
ADRA2C
|
|
Dopamine
|
Dopamine receptor
|
DRD2
|
|
Histamine
|
Histamine receptor
|
HRH1
|
|
HRH2
|
|
HRH3
|
|
5-Hydroxytryptamine
|
Serotonin receptor
|
HTR1A
|
|
HTR2A
|
|
HTR2B
|
|
HTR2C
|
|
Peptide ligands
|
|
Opioids
|
Opioid receptor
|
OPRD1
|
|
Morphine
|
OPRK1
|
|
Endomorphine
|
OPRM1
|
|
Nucleotide ligands
|
|
Adenosine
|
Adenosine receptor
|
ADORA3
|
|
Nucleotides
|
Purinergic receptor
|
P2RY1
|
|
P2RY14
|
|
P2RY4
|
|
P2RY6
|
|
Glucocorticoids
|
|
Cortisol
|
Glucocorticoid receptor
|
NR3C1
|
3.7. Regulation of kinase signaling
External and internal stimuli elicit many signal transduction pathways involved in normal physiological functions of the nervous system. Several pieces of literature have pointed out the dysregulation of different kinases and their regulation in neurodegenerative diseases or neuroprotection. We came across different kinases implicated in neuronal functions whose regulations confer neuroprotection. It is well-known that signaling pathways do not act alone and often result in cross-talks. In the current study, we have mapped some of the pathways, such as MAPK, PI3K-AKT, RAP1, RAS, mTOR, and AMPK, associated with neuronal regulation, which are targets of B. monnieri and C. asiatica metabolites shown in Fig. 6.
3.8. Docking of phytocompounds against target proteins:
We further carried out molecular docking studies to identify potential protein targets of metabolites from B. monnieri and C. asiatica against two selected proteins -A2A adenosine receptor (A2AAR) and monoamine oxidase B (MAO-B). The docking study revealed that in the A2AAR, metabolite Peonidin 3,5-diglucoside showed the highest LibDock score of 175.02 with ten favorable H-bond interactions followed by UDP-B-L-arabinose (164.33), Vicenin-2 (162.755), UDP-alpha-D-sulfoquinovopyranose (158.117), GDP-L-glucose (156.47), respectively. Interestingly we found the phytochemical libraries of both plants docked against A2AAR follow the same docking pattern. Further, we consider the binding site of MAO-B for docking to examine whether any metabolites bind to that site. The results indicated that tetrahydro pteroyl tri-L-glutamate showed the highest docking score of 280.414 with favorable sixteen H-bond interactions and was followed by UDP-alpha-D-sulfoquinovopyranose, Phytofluene, GDP-L-galactose, and dihydrozeatin-9-N-glucoside-O-glucoside with the LibDock score of 225.594, 220.244, 217.511 and 214.563, respectively. Here also, docking studies of both the plant phytochemical libraries showed almost identical order with respect to their docking score. The experimentally determined dual inhibitor 8-(3-chlorostyryl) caffeine (CSC) was used as a standard inhibitor, comparing the docking efficacies of top-ranked phytochemicals. The CSC showed a LibDock score of 105.847 and 123.879 with four and seven H-bond intermolecular interactions, respectively, while docked against A2AAR and MAO-B. This indicates the top-ranked metabolites have more binding affinity than the CSC and can act as dual inhibitors against the targets A2AAR and MAO-B (Fig. 7A-D).
3.9. Validation of neuroprotective activity of B. monnieri and C. asiatica
3.9.1. B. monnieri and C. asiatica aqueous extracts prevent neuronal apoptosis in cellular models of Alzheimer’s and Parkinson’s disease model
IMR-32 cells were differentiated with 10 µM retinoic acid for seven days. They were treated with rotenone (100 nM) to generate the Parkinson’s disease model, 100 µg/ml of B. monnieri and C. asiatica extracts, Aβ₄₂ (0.5µM) to generate the Alzheimer’s disease model, co-treatment of rotenone with B. monnieri or C. asiatica extract, and co-treatment of Aβ₄₂ with B. monnieri or C. asiatica extracts. After drug treatment for 48hrs, cells were subjected to acridine orange - ethidium bromide (AO-EB) dual nuclear staining. The green fluorescence displayed by the control cells indicates its viable nature. The rotenone-treated cells showed early apoptosis with yellowish-green fluorescence, and Aβ42 treated cells showed early and late apoptosis with yellow and light orange fluorescence. Co-treatment with B. monnieri or C. asiatica aqueous extract significantly decreased the cell apoptosis compared to the rotenone and Aβ42 treated cells (Fig. 8A-B), indicating neuroprotective properties of B. monnieri and C. asiatica.
3.9.2. B. monnieri and C. asiatica aqueous extracts prevent excessive reactive oxygen species (ROS) production in cellular models of Alzheimer’s and Parkinson’s disease model
We tested the ROS production by staining the cells with 2′,7′-dichlorofluorescein diacetate (DCFDA). ROS production increased with the rotenone and Aβ42 treatment conditions, which was decreased with the B. monnieri / C. asiatica aqueous extract co-treatment. Rotenone and Aβ42 treatment increased ROS production (54.68% & 70.96% respectively, p < 0.0001) when compared to the control (7.561% (PD) & 9.47% (AD) conditions respectively). On the other hand, co-treatment with B. monnieri / C. asiatica aqueous extract significantly decreased the ROS accumulation when compared to rotenone (22.03% & 21.32%, respectively, p < 0.0001) and Aβ42 (46.60% & 34.72% p < 0.0001 respectively) The results are represented in Fig. 9A-D.
{kind=link}