In the present study, we used data and sera from previous studies to characterize the potential of a BBMA for assessing the serological response of chickens against T. gondii. To our knowledge, this is the first description of a T. gondii-specific serological BBMA for chickens. BBMAs have a number of advantages as compared to MAT, IFAT and ELISA. First, they allow for the simultaneous serological testing of antibodies directed against several pathogens by using a number of antigens coupled to beads with individual colour codes (also called regions). Moreover, with analytes coupled to different bead regions and added to the same BBMA one can implement internal standards, such as a control for sufficient IgY levels in a test sample. Such internal controls can often not be included in other serological assays such as ELISA, IFAT or MAT, when testing individual samples, or only at the expense of testing additional samples.
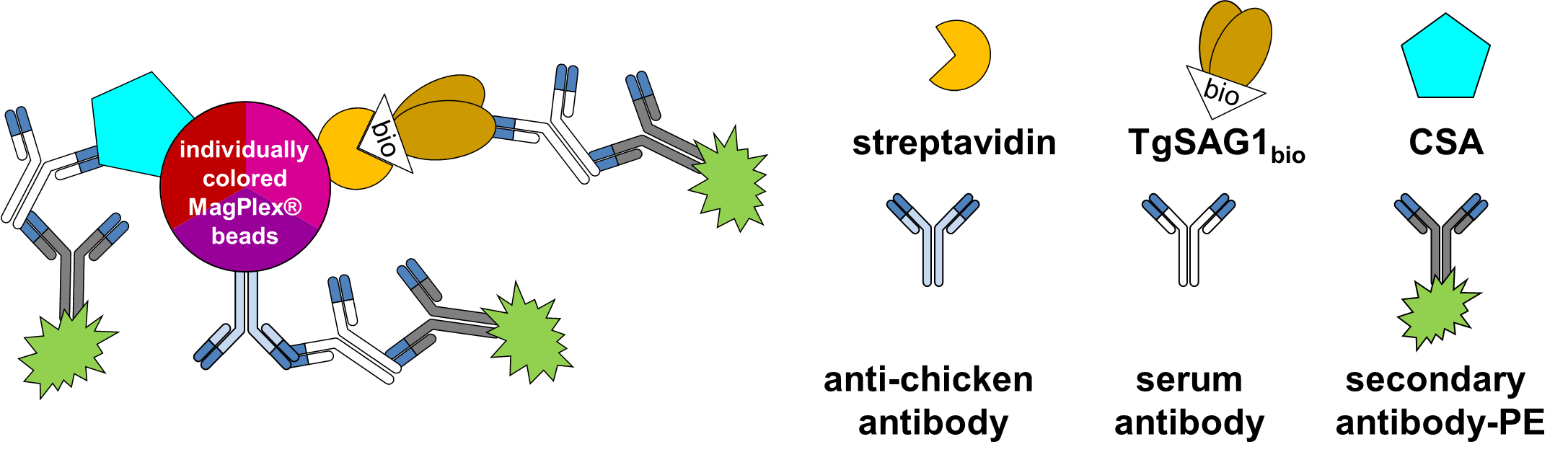
A similar approach for detecting T. gondii infections in animals other than chickens using BBMA has been reported [45, 46]. This assay used a T. gondii tachyzoite lysate as antigen. In contrast, we applied a bacterially expressed biotinylated recombinant TgSAG1 [37], the major surface antigen of T. gondii tachyzoites [24], which is used widely for serodiagnosis in humans, but also in wild or livestock animals (e.g. [47, 48, 49, 50, 51, 52, 53, 54, 55]). Our recombinant TgSAG1bio is unique in that it allows an oriented and reproducible coupling of the antigen to streptavidin-coated magnetic Luminex beads via a single C-terminal biotin [37]. It thus adopts a similar orientation on the beads as native TgSAG1 does on the parasite surface through its GPI anchorage in the membrane [56], thereby exposing the major conformational epitope recognized by human antibodies to the solute [57]. It is not known, if this epitope is also important in chickens.
Only two other studies have reported the use of recombinant TgSAG1 for serodiagnosis in chickens [47, 58]. In both cases, TgSAG1 was used in a denatured form on immunoblots, leading to low sensitivity compared with other recombinant tachyzoite antigens or total lysate, respectively. In contrast, TgSAG1 used in our reference ELISA is purified from tachyzoite lysate by monoclonal antibody affinity chromatography [9]. We conclude that TgSAG1bio as used here is as good as the native protein, but available in larger amounts with less effort.
Since the TgSAG1bio-BBMA was previously tested for human sera and showed excellent diagnostic characteristics in comparison to commercial diagnostic assays [37], we extended the use of the TgSAG1bio-BBMA to chicken sera. We compared the diagnostic characteristics not only with those of other serological tests, but also with the infection status of the birds as determined by samples from experimentally or naturally exposed chickens, by direct detection including conventional PCR on pepsin-digested muscle tissues, MC-qPCR or mouse bioassay.
By using sera of experimentally inoculated chickens, a cut-off was established to separate T. gondii-inoculated from non-infected chickens. With this cut-off, the diagnostic performance of the TgSAG1bio-BBMA in experimental chickens was as good as or superior to that of the ELISAs, IFAT and MAT performed in comparison. Moreover, much higher MFI values were observed in T. gondii inoculated chickens that had tested positive by direct detection (MC-qPCR) than in inoculated chickens without directly detectable infection (Fig. 1). However, we cannot exclude that also inoculated animals without a positive MC-qPCR result were viably infected. Probably, T. gondii had multiplied better in MC-qPCR-positive animals, which may have resulted in a wider distribution of the parasite and could thus have increased the chance to detect the infection by MC-qPCR. At the same time, it may have led to increased exposure to parasitic antigens, including TgSAG1, and thus to a higher level of specific antibodies. In a previous study, it has been observed that higher antibody levels increased the likelihood of a positive result in MC-qPCR in sheep [29].
The cut-off established by using experimental chicken sera proved also suitable in naturally infected chickens. With this cut-off, it was possible to separate sera that had tested positive or negative in a number of reference tests including two ELISAs, IFAT and MAT with high diagnostic sensitivity and specificity. Moreover, especially the TgSAG1-ELISA indices, but also the titres in IFAT or MAT correlated significantly with the MFI values of the TgSAG1bio-BBMA.
In naturally exposed chickens that had also been examined by direct detection methods, the TgSAG1bio-BBMA showed both high diagnostic sensitivity and specificity. The TgSAG1-ELISASL was the only test that had a higher diagnostic sensitivity compared to the TgSAG1bio-BBMA. However, its diagnostic specificity was much lower.
The second cut-off established in experimental chickens (MFI = 3,092) allowed separating birds that were positive in direct detection from birds that were inoculated, but appeared negative in direct detection. In naturally exposed chickens, however, the assay failed to detect 29.0% (9/31) of the direct detection-positive chickens, including eight birds, in which the mouse bioassay had proven a viable T. gondii infection. Therefore, we do not recommend applying this second cut-off.
There is a number of reasons, why the findings obtained with experimentally infected chickens do not necessary match the results for naturally exposed birds: (i) Infections in naturally exposed chickens may date back much further than the more recent infections in experimentally inoculated chickens. Therefore, the exposure to tachyzoite antigens and to TgSAG1 in particular may have occurred more recently in experimentally exposed chickens, which might have had the effect that antibody levels to this antigen are still higher. (ii) Experimentally inoculated chickens may have been exposed to higher parasite doses as compared to naturally exposed chickens. Even low oocyst numbers, which are still infective, but might result in a lower tachyzoite burden, may have induced the development of tissue cysts and eventually viable T. gondii infection in naturally exposed birds. The differences between experimentally and naturally exposed chickens show, that test development and validation in veterinary medicine should never rely on data obtained by experimental infections alone.
{kind=link}