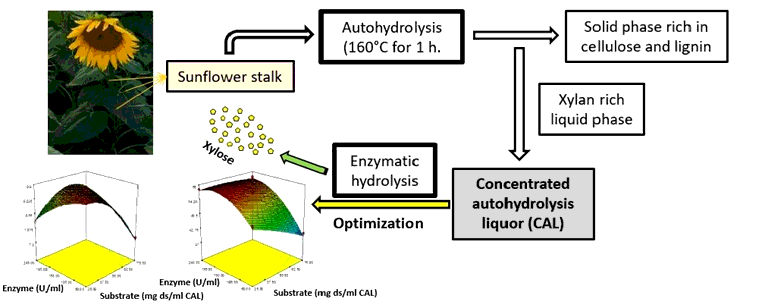
The purpose of this study was to obtain xylose-based fermentation media from autohydrolysis liquors of sunflower stalk by using commercial xylanase formulation. Xylose is generally produced from xylan by diluted acid hydrolysis that causes the formation of some unwanted compounds during the process. As an alternative to dilute acid hydrolysis method, enzymatic hydrolysis of xylan can provide more specific hydrolysis under moderate conditions and does not cause the formation of undesirable compounds. In this study, xylose production carried out with Trichoderma longibrachiatum xylanase on solubilized xylan form of sunflower stalk, which was hydrothermally pretreated for 1 hour at 160ºC. The effects of substrate concentration and enzyme activity were investigated for the production of xylose. To obtain a high xylose yield and selectivity, the optimization study was conducted by the response surface methodology. The optimum substrate concentration and enzyme activity were found as 60 mg ds/mL CAL and 234 U/mL, respectively. Under the optimum condition, xylose yield and selectivity were found to be 69.5% and 8.2 g/g, respectively. This study showed that xylose could be produce with a high yield without requiring a neutralization process and corrosive chemical reagent apart from water.
Research Article
Xylose Release From Sunflower Stalk by Coupling Autohydrolysis and Enzymatic Post-Hydrolysis
https://doi.org/10.21203/rs.3.rs-348348/v1
This work is licensed under a CC BY 4.0 License
Journal Publication
published 22 Oct, 2021
Read the published version in Waste and Biomass Valorization →
You are reading this latest preprint version
Xylose
Xylanase
Autohydrolysis
Sunflower stalk
Optimization
This study presented an alternative route for manufacturing xylose-based fermentation media from sunflower stalk in an environmentally friendly way. Xylose production from sunflower stalk was accomplished by coupling the hydrothermal treatment with the enzymatic post-hydrolysis. The suggested method did not require the neutralization process and did not produce the neutralization products. This study is providing useful information for the future study of the production of xylitol with fermentation and contributing the information for the commercial microbial manufacture of xylitol.
Xylose, a five-carbon pentose sugar, which is the main building block of hemicellulose, is derived from xylan-rich lignocellulosic materials and can be the source for the production of various high-value compounds such as xylitol, sugar alcohol [1,2]. Xylitol is used in many fields such as medicine, food and cosmetics industry due to its low calorie value, insulin independent metabolization and not participating in Maillard reaction with amino acids [1]. The most critical step during the manufacture of xylitol is the hydrolysis of hemicellulosic fraction of lignocellulosic materials to release xylose. This can be achieved by either acid or enzyme catalyzed reactions [3-5].
Agricultural waste consists of three fractions as cellulose, hemicellulose and lignin. The ratio of these fractions varies according to plant species [6]. The hemicellulose structure contains xylan as the main component [7-9]. It consists of β-D-xylopyranose units linked by β-1-4 glycosidic bonds with arabinose, glucuronic acid, methyl and acetyl groups added to the xylose backbone [9, 10]. Due to the amorphous structure of the hemicellulosic fraction of the lignocellulosic biomass [9], it can be hydrolyzed by dilute acid, but its lignin and cellulosic fractions remain unaffected by acid. Although the simplicity and faster kinetics make the acid-catalyzed production of xylose a widely used procedure, the acid causes the equipment corrosion, requires neutralization of the hydrolysate and generates some degradation products acting as a potential fermentation inhibitor for the bioconversion process of xylose to other products [5]. These toxic compounds include furan, aliphatic acids and phenolic compounds [4].
The use of xylanase to hydrolyze xylan may be another alternative to acid hydrolysis of it for the release of xylose [3, 8]. Unlike acid hydrolysis, enzymatic hydrolysis takes place in milder conditions, does not require corrosive chemical or create industrial waste and degradation products. Since the procedure is more specific method, it allows better fermentability of the xylose solution by the microorganism [5, 11]. The enzymatic method of saccharification hemicellulose is specific to xylose and does not result in microbial inhibitors [3].
Since xylan forms a complex with lignin that makes lignocellulosic biomass resistant to acid or enzymatic hydrolysis, xylose production carries out with two steps: pretreatment of lignocellulosic biomass and hydrolysis either acid or enzyme of it [12, 13]. For the enzymatic hydrolysis of hemicellulose, lignocellulosic biomass can be treated with a chemical, mechanical, hydrothermal and/or a combination of these methods [5, 7, 9, 14, 15]. Among them, hydrothermal pretreatment, known as autohydrolysis, has proven to be an effective method for the solubilization of hemicellulose to oligomers [9, 16-19]. When xylan including lignocellulosic biomass is treated at high temperatures (140-220 ℃) for a definite time, the hydronium ions come from water autoionization and acidic groups of xylan depolymerize and solubilize it [9, 20]. Cellulose and lignin are not significantly affected by the hydrothermal pretreatment [21]. Since water is used during the process of hydrolysis, a solid phase rich in cellulose and lignin and a liquid phase containing a low concentration of undesirable by-products (furfural or lignin hydrolysis products) are obtained as the hydrolysis products [3, 11, 21].
Sunflower, grown in temperate and subtropical climates, is one of the world's major oil seeds and the crop's turkey [22]. It is the third oilseeds produced source [23]. that is cultivated in 27,37 million of the world land area [24]. Based on the 2019 FAO statistical data, Turkey is 6th in the world with 2,1 million tons of sunflower production [24]. The stalks of the sunflower plant, whose seeds are harvested, can be used as raw materials for the production of paper pulp and bioethanol [25], but these uses represent a small portion of them. Mainly, they are left behind in the fields to rot or burned in the fields that are associated with environmental risks [26]. In Turkey, around 2.5 million tons of waste per year in the form of sunflower stalks are produced [27], and this biomass can be an abundant and renewable resource for soluble sugar, especially xylose. The previous study indicated that xylan obtained from sunflower stalk has a more linear structure than softwood xylan and consists of primarily xylose units (> 90%) [28].
The release of sugars from the lignocellulosic wastes by coupling hydrothermal treatment and enzymatic post-hydrolysis has been studied by some researchers. However, most of these studies reported enzymatic hydrolysis of solids spent rather than liquids of lignocellulosic materials [3, 17, 29-32]. The previous studies were done on corn cob and corn bran autohydrolysis liquors showed that xylose can be obtained with 80% and 36% yield, respectively [5,33]. Another study reported that autohydrolysis of corn bran and its subsequent enzymatic hydrolysis revealed that the xylose yield was more than 80% [16].
This study aimed to evaluate the commercial xylanase formulation for obtaining xylose-based fermentation media from autohydrolysis liquors of sunflower stalk. Based thorough our literature review, we have not coincided with a study about xylose production from sunflower stalk by coupling the hydrothermal treatment with the enzymatic post-hydrolysis. It was reported that xylanase from the GH10 family produces more monosaccharides than xylanase from GH11 family [12]. In this research, Trichoderma longibractium xylanase from GH10 family was preferred for the post-hydrolysis for liquors obtained from the autohydrolysis. This study determined the optimum substrate concentration and enzyme activity by using response surface methodology for the production of xylose with high yield.
2.1. Materials
Sunflower stalks gathered from local farmers in Turkey, dried with air at room temperature up to 8.3% moisture content and ground to particles of 2-10 mm in length. A commercial endo-xylanase, from Trichoderma longibrachiatum was obtained from Danisco (Finland) and Aminex HPX 87H column (dimension: 300x7.8 mm; average particle size: 25 mm) was purchased from Bio-Rad Laboratories (CA, USA). All other chemicals used in this study were from Sigma Chemical Company (MO, USA) or Merck KGaA(Germany) or Alfa Aesar GmbH & Co KG (Germany).
2.2. Autohydrolysis
Sunflower stalk was mixed with water in a ratio of 8:1 kg/kg (liquid/solid) in the batch 1 L- stainless steel reactor (Parr Reactor, USA). Its autohydrolysis took place at 160°C for 1 h. Spent solid from autohydrolyzed sunflower stalk (SSAS) was recovered by the filtration, washed with water and air-dried, and its moisture was determined to measure the amount of the dissolved substrate (ds). The filtrate was concentrated by vacuum evaporation below 50oC and called concentrated autohydrolysis liquor (CAL).
2.3. Analysis of concentrated autohydrolysis liquor and spent solid from autohydrolysis
Moisture, ash and lignin (Klason and acid soluble) content of SSAS were determined according to the ASTM [34]. The polysaccharides of the SSAS were hydrolyzed and their monosaccharides compositions were determined [35, 36]. Ground SSAS (300 mg) was mixed with 72% (w/w) sulphuric acid (3 mL), held at 30°C for 1 h with stirring and at the end of 1 h, acid concentration was diluted to 4% (g/g) with water. After refluxing the mixture for 4 h, the sugars in the aliquot of the hydrolysate were analyzed by HPLC. The amount of polysaccharides and acetyl groups in the reaction products were determined by measuring the monosaccharides and acetic acid concentration by HPLC and converted to percent parent polysaccharides (D-glucose to cellulose, D-xylose to xylan and D-arabinose to arabinan). Uronic acid and reducing sugar contents of SSAS were determined by the m-hydroxydiphenyl method using glucuronic acid as the standard [37] and DNS method [38] using xylose as the standard, respectively. Furfural amount in the liquor was determined by adding 1 volume of ethanol into the sample and its absorbance was measured at 277 nm [39].
HPLC analysis of the sample carried out on a system that had a refractive index detector (Perkin Elmer Series 200), column oven (Perkin Elmer Series 200) and Aminex HPX 87H column. The filtered (0.20 mm membranes) CAL and hydrolyzed CAL with 4% (g/g) sulfuric acid were injected into the HPLC, and the monosaccharides and acetic acid were eluted from the column at 45oC by using 5 mmol/L H2SO4 as the mobile phase (flow rate of 0.5 mL/min). The oligomer concentration was determined by the increase in the concentrations of monosaccharides before and after the hydrolysis [40].
2.4. Enzymatic xylan hydrolysis
The enzyme activity of T. longibrachiatum xylanase was determined by endo-xylanase assay by using birchwood xylan as a substrate and one unit is defined as the quantity of the enzyme that liberates 1.0 mmol of xylose per minute from the substrate [41]. Hydrolysis of CAL, 10 mL pH 4.6, was started by the mixing of 1 mL of (23-278 U/mL) xylanase from T. longibrachiatum at 50oC. At regular time intervals, 1 mL sample was taken and boiled at 100oC for 5 min to inactivate the enzyme and analyzed by its reducing sugar and HPLC as described above.
2.5. Experimental design and response surface methodology
A 22 central composite design (CCD), consisted of 13 experiments, was used for the optimization study. Substrate concentration (14.65 and 85.35 mg ds/mL CAL) and enzyme activity (22-277 U/mL) were used as independent variables (Table 1) while xylose yield and selectivity were taken as the dependent variables. The quadratic model was selected for the predicting of the optimal point and expressed as

where Y represents response variables (xylose yield and selectivity); b0 is the interception coefficient; b1 and b2 are the linear terms; b11 and b22 are the quadratic terms; b12 is the coefficient for the interaction between X1 (substrate concentration) and X2 (enzyme activity) that represent the variables studied.
The Design Expert v.7 (Stat-Ease Inc., Minneapolis) program was used for the regression and graphical analyses of the data obtained. The equations of both models and their statistical significances of the regression coefficients were determined by the Fischer's test and Student’s t-test, respectively. The graphical optimization to find the optimum concentrations of the substrate and enzyme was conducted using the Design-Expert program.
3.1. Composition of autohydrolysis liquor and spent solid of autohydrolyzed sunflower stalks
In the former work, it was reported that cellulose (36%) was the main component of sunflower stalk, followed by xylan (22%) and lignin (klason lignin: 26% and acid soluble lignin: 1%). The rest of the constituents of it were arabinan (0.7%), acetyl groups (3%), uronic acid (6%), protein (1%) and ash (3%) [28].
The autohydrolysis of sunflower stalk was accomplished at 160oC for 1 h according to the result of the previous study [12] and from the autohydrolysis of 100 kg sunflower stalk, 25 kg of the total dissolved substrate (ds) was obtained. The main components of the liquor were xylooligomers, some monosaccharides (mainly xylose followed by glucose and arabinose) and acetic acid, which were derived from either cellulose or hemicellulose fraction of the sunflower stalk. The resulting liquor was concentrated and the composition of concentrated autohydrolysis liquor (CAL) is presented in Table 2.
The concentration of xylooligomer in the CAL of the sunflower stalk was 27 g/L. In addition to xylooligomer, acetyl groups linked to oligomer, glucooligomer deriving from cellulosic components of the raw materials, arabinooligomer were present in the CAL, and their levels were 3.30, 1.35 and 1.50 g/L, respectively. The furfural production, dehydration product of the sugars, remained at low levels (Table 2) when the results were compared with a previous study done with dilute acid hydrolysis [28].
As expected, the pretreatment solubilized the hemicellulosic fraction of the sunflower stalk to a certain grade and changed the combination of dry matter. Table 3 indicates the composition of spent solid from autohydrolyzed sunflower stalk (SSAS). With the hydrolytic treatment of lignocellulosic materials, hemicellulose became partially soluble, the main component of SSAS was cellulose (43%) followed by lignin (32%) and hemicellulose (18%). The previous studies on the determination of the composition of spent solid of the hydrothermally treated sunflower stalk showed similar results, its major component was cellulose (47.9%) followed by lignin and hemicelluloses [32, 42].
The different studies reported similar results for the autohydrolysis of other lignocellulosic material sources. The autohydrolysis of agave bagasse at 160 ℃ for 50 min showed that it had a main component as cellulose (35%), followed by lignin (25%) and hemicellulose (6%) [31]. The previous study presented that cellulose (78%), lignin (6%) and hemicellulose (1%) constituted the solid phase of autohydrolyzed wheat straw (at 190℃ for 15 minutes) [43]. It was reported that the xylan content in the solid residues decreases with the increase of the pretreatment temperature [17, 19]. The differences in composition and structure of the sunflower stalk and autohydrolysis process conditions were thought to be the reasons for differences in results between this study and those previously reported.
3.2. Enzymatic hydrolysis
To get a loud yield of xylose, CAL was hydrolyzed by T. longibrachiatum xylanase from the GH10 family, the commercial endo-xylanase preparation. In general, unlike GH11 xylanases, GH10 xylanases produce low-DP oligosaccharides, mainly xylose monomer and xylobiose [44, 45]. Since the autohydrolysis solubilizes the hemicellulosic fraction of the sunflower stalk as oligomers, its CAL is an ideal substrate for the xylanases to produce xylose.
The use of the high concentration of the substrate may decrease the yield due to enzyme inhibitory activity that presents in the reaction mixture, while keeping the enzyme level high may cause an increase in product cost. Therefore, it is necessary to find the ideal substrate concentration and enzyme activity. The different concentrations of CAL of the sunflower stalk were treated at different levels of T.longibrachiatum xylanase for 46 hr at pH 4.6 and 50 oC. The experimental range and levels of the substrate concentration and enzyme activity investigated are given in Table 1. It was observed that all the hydrolysis progress curves of CAL (Figure 1) showed an increase in the first 24-hr and flatten after 24 hr. The highest reducing sugar was obtained at 11th run (75 mg ds/mL CAL and 240 U/mL) and the reducing sugar concentration of this run was 21 mg/mL at the end of the 24 hr. The lowest reducing sugar was obtained at the 10th run (14.65 mg ds/mL CAL and 150 U/mL) and it was 3.8 mg/mL after 24 hr hydrolysis period.
The compositions of the hydrolysis products at the end of 24-hr reaction are presented in Figure 2. The highest xylose (10.2 mg/mL) and glucose (1.1 mg/mL) were found at 11th run and the highest arabinose concentration (0.022 g/L) was found in the 3rd run. Since plant xylans are partially acetylated, the concentration of acetic acid increased with the increase in the substrate concentration. The highest acetic acid (1.9 mg/mL) concentration was found in the 13th run.
Since the solid residue, SSAS, had 18% hemicelluloses (Table 3), it was also hydrolyzed with T. longibrachiatum xylanase for 48 hr at pH 4.6 and temperature 50 °C. The result of the hydrolysis reaction (Figure 3A and B) was monitored by measuring the individual sugar concentration in the hydrolysate and the highest sugar concentration was determined as xylose and then glucose. The highest xylose yield was found after 24-hr hydrolysis period (19% or 3.0 mg/mL xylose) while the highest glucose level was found after the 48-hr hydrolysis period (2.5 mg/mL) (Figure 3A). The conversion of residual xylan to xylose increased with the hydrolysis time, but the selectivity (the ratio of xylose to other sugar) decreased due to the release of glucose and arabinose (Figure 3B). This is because xylanases from the GH10 family are less specific to xylan [8] and can also act on cellulose substrates [46].
It is not easy to compare the results we obtained in this study with the results obtained in previous studies due to differences in the type of biomass or hydrolytic reagent. The spent solid is mainly composed of cellulose followed by xylan, if cellulase was used as an enzyme, the yield of glucose is higher or if xylanase was used, the yield of xylose is higher at the end of the hydrolysis process. A study on the enzymatic hydrolysis of spent solid of hydrothermally processed corn bran with different xylanase activity from Eupenicillium parvum 4-14 and Aspergillus oryzae showed that A. oryzae derived enzyme had higher cellulase activity and it produced xylose and glucose with 6.96% and 31.04% yields, while E. parvum 4-14 derived enzyme had high hemicellulase activity and produced xylose and glucose with 45.43% and 16.77% yields [16]. The study done on the spent solid of the hydrothermally treated corn bran with enzymes with xylanase different commercial xylanase showed that xylose and glucose were obtained as 8% and 1% yield, respectively, but when cellulase was used for the enzymatic hydrolysis, glucose yield was more than 25% [3]. The previous studies on the production of sugars from enzymatic hydrolysis of spent solid of hydrothermally treated lignocellulosic biomass showed that cellulosic enzymes produced glucose with a higher yield than hemicellulosic sugars [3, 16, 29, 32].
3.3. Optimization
The design of this research including response variables is given in Table 4, the percentage conversion ratio of xyloligomers to xylose in autohydrolysis liquors is expressed as xylose yield, the ability of xylanase to hydrolyze xylan is expressed as selectivity.
In this study, high xylose yield and selectivity were desired to keep other hydrolysis products low in the hydrolyzate. According to Table 4, the highest xylose yield is 59.09% (substrate: 50 mg ds/mL CAL, enzyme: 150 U/mL), the highest selectivity is 9.90 g/g, obtained, at run 9 (substrate: 25 mg ds/mL CAL, enzyme: 60 U/mL). The lowest xylose yield is 37.42% at run 7 (substrate: 50 mg ds/mL CAL, enzyme: 22.74 U/mL). Among the experiments with the same substrate concentrations, but different enzyme activity (runs 1, 2, 5, 6, 7, 8 and 12), run 7 was found to have the lowest xylose yield. At the same substrate concentration, increasing enzyme level from 22.74 to 150 U/mL increased the xylose yield. However, increasing the enzyme level to more than 150 U/mL did not make significant changes in the yield.
The quadratic models for xylose yield and selectivity are shown in the Eq. (2) and Eq. (3), in which Y1 and Y2 represent the xylose yield and selectivity, respectively, as the function of the substrate level (X1) and enzyme activity (X2).
To fit the experimental data and response function, the regression analysis was performed. The quadratic models for xylose yield and selectivity were appraised by ANOVA (Table 5). The p-values (<0.05) and F values (33.4 and 102.44) of both models showed their significance. Besides, the models did not show a lack of fit and the determination coefficients (R2) for the xylose yield (Y1) and the selectivity (Y2) were 0.96 and 0.99, respectively. The adjusted determination coefficient (Adj R2, 0.93 and 0.98) also verified the importance of the models. Table 5 presents Pred R2 values that are in acceptable accord with the Adj R2 values, showing a good adjustment between the determined and estimated values. It was seen that the experiments were precise and reliable, with lower values of the coefficient of variation (C.V. 3.41 and 1.34%). Adequate precision (signal to noise ratio) greater than 4 indicates adequacy of the model precision. The ratios of both the models (16.7 and 30.3) were found greater than 4 (Table 5).
Figure 4 presents the diagnostic plots to be used to assess the sufficiency of the models. As seen from Figures 4A and 4D, the predicted and determined values for both responses are in agreement with each other sufficiently. A normally distributed was observed in the normal % probability plots of residuals for both responses without deviation of the (Figure 4B and 4E). All the data points lied within the limits (±3) in the internally studentized residuals plots (Figure 4C and 4F).
Figure 5 presents the response surface plots to predict the xylose yield and selectivity over the independent variables (substrate concentration and enzyme activity). The maximum xylose yield (59%) was obtained at 36 mg ds/mL CAL substrate level and 196 U/mL enzyme activity (Figure 5A). The maximum selectivity (9.8 gg-1) was get at 25 mg ds/mL CAL substrate concentration and 60 U/mL enzyme activity (Figure 5B).
Decreased xylose yield and increased selectivity with the increase in the substrate concentration, while increased xylose yield and decreased selectivity with the increase in enzyme level were observed (Figure 2 and 5). Both the factors displayed (enzyme and substrate level) significant linear (p<0.05) effects on xylose yield. Substrate concentrationxenyzme level showed positive significant effects on both responses. The quadratic coefficient of enzyme level presented significant negative effects on both responses, but that of substrate concentration showed significant effect on the selectivity and non significant effect on the yield (Table 5).
Based on the two models, a graphical optimization was conducted by overlaying the contour plots of the models (Figure 6). The optimum operating condition with high levels of xylose efficiency and selectivity was chosen by applying the following criteria: xylose yield>50 and selectivity> 9 gg-1, and 60 mg ds/mL CAL substrate concentration and 234 U/mL enzyme level were chosen as the optimal working conditions foreboded through the program. To confirm the result, the enzymatic hydrolysis was conducted in duplicate at the optimum conditions. The change in xylose, glucose, acetic acid, xylose yield and selectivity along the hydrolysis process, conducted at the optimum conditions, are offered in Figure 7.
As appeared from Figure 7A, xylose concentration increases up to 24 hr. The maximum xylose concentration was measured at 48 hr as 9.3 mg/mL; after this point, its concentration decreased. The maximum glucose concentration was measured at 72 hr as 1.7 mg/mL. Xylose yield was found as 70% for 24 hr and 72% for the 48 hr hydrolysis period while selectivity was found maximum at 24 hr as 8.2 g/g and decreased with time (Figure 7B). Since the reaction taking longer than 24 hr did not cause an important rise in xylose yield, 24-hr hydrolysis seems suitable for the production of xylose.
Table 6 presents the results of the optimization work and the results of estimated conditions suggested by the statistical program used for the optimization. Xylose yield obtained in the experiment is higher than the predicted value while selectivity is slightly lower than the predicted value.
The differences in lignocellulosic material, pretreatment conditions and enzyme source make it difficult to compare the data obtained in this study with the results obtained from previous studies. In the previous study about the enzymatic hydrolysis of palm waste autohydrolysis liquor (treated at 121ºC for 20-80 min) with T. viride xylanase, xylose yield was found as 25.64% [47]. Another study about hydrolysis of the autohydrolysis liquor of corn husk, obtained at 190ºC for 10 min, with the enzyme cocktail including endoxylanase, beta xylobiase and arabinofuranosidase reported the xylose yield as 35% [37]. A study on the production of xylose by the coupling of the hydrothermal treatment with the enzymatic post-hydrolysis of corn cob found the xylose yield as 80% [5]. As a result of hydrolysis of the liquor obtained as a result of autohydrolysis of tobacco stalks at 160 ° C for 1 hour, 79.8% xylose yield was obtained with T. longibrachiatum xylanase [48]. The hydrolysis of the autohydrolysis liquor of corn bran, obtained at 165ºC for 40 min, with the enzyme blend from Aspergillus oryzae and Eupenicillium parvum xylanase reported the xylose yield as more than 80% [16]. The results obtained from this work seen that the xylose can be obtained with high yield by the coupling of the hydrothermal treatment and enzymatic hydrolysis methods.
This study presented an alternative way to produce xylose from sunflower stalk by combining enzymatic hydrolysis and autohydrolysis. The optimum substrate and enzyme concentration were found as 60 mg ds/mL CAL and 234 U/mL, respectively where xylose yield and selectivity were 69.5% and 8.2 g/g. This research provides information about the enzymatic production of xylose with a high yield without requiring a neutralization process and corrosive chemical reagent apart from water. Besides, this study supplies beneficial data for further works on the manufacture of xylitol with fermentation and contribute to the information for the commercial microbial production of xylitol.
Acknowledgments
This work was financially supported by Gaziosmanpasa University Research Fund (2013/46).
Statement of Novelty
This study presented an alternative route for manufacturing xylose-based fermentation media from sunflower stalk in an environmentally friendly way. Xylose production from sunflower stalk was accomplished by coupling the hydrothermal treatment with the enzymatic post-hydrolysis. The suggested method did not require the neutralization process and did not produce the neutralization products. This study is providing useful information for the future study of the production of xylitol with fermentation and contributing the information for the commercial microbial manufacture of xylitol.
Sample Credit author statement
Title: Optimization of Enzymatic Processing Conditions for Xylose Release from Sunflower Stalk Autohydrolysis Liquor
Author's contribution:
Fatmagül Halici Demir: Methodology, Investigation, Data analysis, Writing manuscript
Özlem Akpinar: Supervision, Reviewing.
- Walsh, M.K., Khliaf, H.F., Shakir, K.A.: Production of Xylitol from Agricultural Waste by Enzymatic Methods. American Journal of Agricultural and Biological Sciences. (2018). https://doi.org/10.3844/ajabssp.2018.1.8
- Rafiqul, S.M., Sakinah, A.M.M., Karim, M.R.: Production of Xylose from Meranti Wood Sawdust by Dilute Acid Hydrolysis. Appl. Biochem. Biotechnol. (2014). https://doi.org/10.1007/s12010-014-1059-z
- Liavoga, A.B., Bian, Y., Seib, P.A.: Release of D-xylose from wheat strawby acid and xylanase hydrolysis and purification of xylitol. J. Agric. Food Chem. (2007). https://doi.org/10.1021/jf070862k
- Mohamad, N.L., Kamal, S.M.M., Mokhtar, M.N.: Xylitol bioproduction: A review of recent studies. Food Rev. Int. (2015). https://doi.org/10.1080/87559129.2014.961077
- Vazquez, M.J., Alonso, J.L., Donminguez, H., Parajo, J.C.: Production of xylose-containing fermentation media by enzymatic post-hydrolysis of oligomers produced by corn cob autohydrolysis. World J. Microb. Biotechnol. 17, 817–822 (2001)
- Bajpai, P.: Structure of Lignocellulosic Biomass. In: Pretreatment of Lignocellulosic Biomass for Biofuel Production. In: Pratima, B. (Ed.), pp. 7–12. Springer: (2016)
- Arora, A., Nandal, P., Singh, J., Verma, M.L.: Nanobiotechnological advancements in lignocellulosic biomass pretreatment. Materials Science for Energy Technologies. (2020). https://doi.org/10.1016/j.mset.2019.12.003
- Moreira, L.R.S., Filho, E.X.F.: Insights into the mechanism of enziymatic hydrolysis of xylan. Appl. Microbiol. Biotechnol. (2016). https://doi.org/10.1007/s00253-016-7555-z
- Sun, S., Sun, S., Cao, X., Sun, R.: The role of pretreatment in improving the enzymatic hydrolysis of lignocellulosic materials. Biores. Technol. (2016). http://dx.doi.org/10.1016/j.biortech.2015.08.061
- Naidu, D.S., Hlangothi, S.P., John, M.J.: Bio-based products from xylan: A review. Carbohyd. Polym. (2018). https://doi.org/10.1016/j.carbpol.2017.09.064
- Rivas, B., Dominguez, J.M., Domingues, H., Parajo, J.C.: 2002. Bioconversion of posthydrolysed autohydrolysis liquors: an alternative for xylitol production from corn cobs. Enzyme Microb. Technol. (2002). https://doi.org/10.1016/S0141-0229(02)00098-4
- Akpinar, O., Gunay, K., Yilmaz, Y., Levent, O., Bostanci, S.: 2010. Enzymatic processing and antioxidant activity of agricultural waste autohydroysis liquors. Bioresour. 5, 699–711 (2010)
- Wan Azelee, N.I.W., Jahim, J.M., Ismail, A.F., Fuzi, S.F.Z.M., Rahman, R.A., Ghazali, N.F., Illias, R.M.: Enzymatic Hydrolysis of Pretreated Kenaf using a Recombinant Xylanase: Effects of Reaction Conditions for Optimum Hemicellulose Hydrolysis. American Journal of Agricultural and Biological Sciences (2016). https://doi.org/10.3844/ajabssp.2016.54.66
- Barberousse, H., Kamoun, A., Chaabouni, M., Giet, J.M., Roiseux, O., Paquot, M., Deroannea, C., Bleckera, C.: Optimization of enzymatic extraction of ferulic acid from wheat bran, using response surface methodology and characterization of the resulting fractions. J. Sci. Food. Agric. (2009). https://doi.org/10.1002/jsfa.3630
- Saha, B.C.: Hemicellulose bioconversion. J. Ind. Microbiol. Biotechnol. (2003). https://doi.org/10.1007/s10295-003-0049-x
- Jiang, K., Li, L., Long, L., Ding, S.: Comprehensive evaluation of combining hydrothermal pretreatment (autohydrolysis) with enzymatic hydrolysis for efficient release of monosaccharides and ferulic acid from corn bran. Industrial Crops & Products (2018). https://doi.org/10.1016/j.indcrop.2018.01.047
- Khalili, F., Amiri, H.: Integrated processes for production of cellulosic and hemicellulosic biobutanol from sweet sorghum bagasse using autohydrolysis. Ind. Crops Prod. (2019). https://doi.org/10.1016/j.indcrop.2019.111918
- Ruiz, H.A., Rodríguez-Jasso, R.M., Fernandes, B.D., Vicente, A.A., Teixeira, J.A.: Hydrothermal processing, as an alternative for upgrading agriculture residues and marine biomass according to the biorefinery concept: A review. Renew. Sustain. Energy Rev. (2013). https://doi.org/10.1016/j.rser.2012.11.069
- Zhang, W., Zhang, X., Lei, F., Jiang, J.: Co-production bioethanol and xylooligosaccharides from sugarcane bagasse via autohydrolysis pretreatment. Renewable Energy (2020). https://doi.org/10.1016/j.renene.2020.10.034
- Gandla, M.L., Martın, C., Jönsson, L.J.: 2018. Analytical Enzymatic Saccharification of Lignocellulosic Biomass for Conversion to Biofuels and Bio-Based Chemicals. Energies (2018). https://doi.org/10.3390/en11112936
- Moniz, P., Ho, A.L., Duarte, L.C., Kolida, S., Rastall, R.A., Pereira, H., Carvalheiro, F.: Assessment of the bifidogenic effect of substituted xylo-oligosaccharides obtained from corn straw. Carbohyd. Polym. (2016). https://doi.org/10.1016/j.carbpol.2015.09.046
- Konyali, S.: Sunflower Production, Consumption, Foreign Trade and Agricultural Policies in Turkey. Social Sciences Research Journal 6(4), 11–19 (2017)
- Anonymous: https://www.statista.com/statistics/267271/worldwide-oilseed-production-since-2008/ (2020). Accessed 28 december 2020
- FAO, 2019. www. faostat.org Accessed 28 december 2020
- Kim, H.S., Oh, Y.H., Jang, Y.-A., Kang, K.H., David, Y., Yu, J.H., Song, B.K., Choi, J., Chang, Y.K., Joo, J.C., Park, S.J.: Recombinant Ralstoniaeutropha engineered to utilize xylose and its use for the production of poly(3-hydroxybutyrate) from sunflower stalk hydrolysate solution. Microb Cell Fact. (2016). doi:10.1186/s12934-016-0495-6
- Sharma, S.K., Kalra, K.L., Grewal, H.S.: Fermentation of enzymatically saccharified sunflower stalks for ethanol production and its scale up. Biores. Technol. (2002). https://doi.org/10.1016/S0960-8524(02)00076-7
- Binici, H., Aksogan, O., Dıncer, A., Luga, E., Eken, M., Isikaltun, O.: Thermal insulation material production using vermiculite, sunflower stalk and wheat stalk. Thermal Science and Engineering Progress (2020). https://doi.org/10.1016/j.tsep.2020.100567
- Akpinar, O., Levent, O., Sabanci, S., Uysal, R.S., Sapci, B.: Optimization and comparison of dilute acid pretreatment of selected agricultural residues for recovery of xylose. BioResources. 6, 4103–4116 (2011)
- Amiri, H., Karimi, K.: Autohydrolysis: A promising pretreatment for the improvement of acetone, butanol, and ethanol production from woody materials. Chem. Eng. Sci. (2015). https://doi.org/10.1016/j.ces.2015.07.020
- Eom, I.-Y., Yu, J.-H.: Structural characterization of the solid residue produced by hydrothermal treatment of sunflower stalks and subsequent enzymatic hydrolysis. J. Ind. Eng. Chem. (2015). https://doi.org/10.1016/j.jiec.2014.07.044
- Pino, M.S., Rodriguez-Jasso, R.M., Michelin, M., Ruiz, H.A.: Enhancement and modeling of enzymatic hydrolysis on cellulose from agavebagasse hydrothermally pretreated in a horizontal bioreactor. Carbohyd. Polym. (2019). https://doi.org/10.1016/j.carbpol.2019.01.111
- Ruiz, E., Cara, C., Manzanares, P., Ballesteros, M., Castro, E.: Evaluation of steam explosion pre-treatment for enzymatic hydrolysis of sunflower stalks. Enz. Microb. Technol. (2008). https://doi.org/10.1016/j.enzmictec.2007.09.002
- Agger, J., Vikso-Nielsen, A., Meyer, A.S.: Enzymatic xylose release from pretreated corn bran arabinoxylan: differential effects of deacetylation and deferuloylation on insoluble and soluble substrate fractions. J. Agr. Food. Chem. (2010). https://doi.org/10.1021/jf100633f
- Phidelphia, P.A.: ASTM, Annual Book of ASTM Standards, American Society for Testing and Materials (04.09). (1993)
- Browning, L.: Methods of Wood Chemistry. Inter-Science Publishers, New York (1967)
- Templeton, D.: Ehrman, T.: Determination of Acid- Insoluble Lignin in Biomass,” Laboratory Analytical Procedure No. 003, National Renewable Energy Labora- tory, Golden, CO (1995)
- Melton, L.D., Smith, B.G.:. Wrolstad, R.E., Acree, T.E., Decker, E.A., Penner, M.H., Reid, D.S., Schwartz, S.J., Shoemaker, C.F., Smith, D., Sporns, P.: Determination of the uronic acid content of plant cell walls using a colorimetric assay. Ed. John Wiley & Sons, Current Protocols in Food Analytical Chemistry, Inc., New York (2001). DOI:10.1002/0471142913.fae0303s00
- Miller, G.L.: Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 31, 426–428 (1959)
- William, S.: AOAC official methods of analysis (9.097), Arlington, VA: (1997)
- Garrote, G., Domınguez, H., Parajo, J.C.: Mild autohydrolysis: an environmentally friendly technology for xyloolgosaccharide production from wood. J. Chem. Biotechnol. (1999). https://doi.org/10.1002/(SICI)1097-4660(199911)74:11<1101::AID-JCTB146>3.0.CO;2-M
- Bailey, M.J., Biely, P., Poutanen, K.: Interlabratory testing of methods for assay of xylanase activity. J. Biotechnol. (1992). https://doi.org/10.1016/0168-1656(92)90074-J
- Hesami, S.M., Zilouei, H., Karimi, K., Asadinezhad, A.: Enhanced biogas production from sunflower stalks using hydrothermal and organosolv pretreatment. Ind. Crops Prod. (2015). https://doi.org/10.1016/j.indcrop.2015.07.018
- Bassani, A., Fiorentini, C., Vellingiri, V., Moncalvo, A., Spigno, G.: Implementation of Auto-Hydrolysis Process for the Recovery of Antioxidants and Cellulose from Wheat Straw. Appl. Sci. (2020). https://doi.org/10.3390/app10176112
- Beaugrand, J., Chambat, G., Wong, V.W., Goubet, F., Rémond, C., Paes, G.: Impact and efficiency of GH10 and GH11 thermostable endoxylanases on wheat bran and alkali-extractable arabinoxylans. Carbohydrate Res. (2004). https://doi.org/10.1016/j.carres.2004.08.012
- Rahmani, N., Kahar, P., Lisdiyanti, P., Jaemin, L., Yopi, Prasetya, B., Ogino, C., Kondo, A.: GH-10 and GH-11 Endo-1,4-β-xylanase enzymes from Kitasatospora sp. produce xylose and xylooligosaccharides from sugarcane bagasse with no xylose inhibition. Bioresour. Technol. (2018). https://doi.org/10.1016/j.biortech.2018.10.00
- Collins, T., Gerday, C., Feller, G.: Xylanases, xylanase families and extremophilic xylanases. FEMS Microbiol. Rev. (2005). https://doi.org/10.1016/j.femsre.2004.06.005
- Sabiha- Hanım, S., Noor, M.A.M., Rosma, A.: Effect of autohydrolysis and enzymatic treatment on oil palm (Elaeisguineensis Jacq.) frond fibres for xylose and xylooligosaccharides production. Bioresour. Technol. (2011). https://doi.org/10.1016/j.biortech.2010.08.017
- Halici Demir, F., Akpinar, O.: Tütün Saplarından Kombine Otohidroliz ve Enzimatik Hidroliz ile Ksiloz Üretimi. Akademik Gıda (2018). https://doi.org/10.24323/akademik-gida.415613
Table 1. Experimental range and levels of the independent process variables
|
Independent variable |
Symbol |
Range and levels |
||||
|
-a |
-1 |
0 |
+1 |
+a |
||
|
Substrate concentration (mg ds/mL CAL) |
X1 |
14.65 |
25 |
50 |
75 |
85.35 |
|
Enzyme activity (U/mL) |
X2 |
2.27 |
6 |
15 |
24 |
27.73 |
Table 2. Composition of the concentrated autohydrolysis liguors (CAL) of sunflower stalk
|
Compound |
Amount (g/L concentrated autohydrolysis liquor) |
|
Glucose (g/L) |
1.90±0.30 |
|
Xylose (g/L) |
5.00±0.09 |
|
Arabinose (g/L) |
0.45±0.02 |
|
Acetic acid (g/L) |
1.72+0.15 |
|
Glucooligomer (g/L) |
1.35±0.15 |
|
Xylooligomer (g/L) |
26.58±4.72 |
|
Arabinooligomer (g/L) |
1.50±0.10 |
|
Acetyl groups linked to oligomer (g/L) |
3.30±0.42 |
|
Furfural (g/L) |
0.64±0.04 |
|
Uronic acid (g/L) |
0.05±0.00 |
Table 3. Composition of the spent solid from autohydrolyzed sunflower stalk (SSAS)
|
Compound |
Amount (g/100 g SSAS) |
|
Cellulose |
42.62±1.4 |
|
Xylan |
18.07±0.93 |
|
Arabinan |
0 |
|
Acetyl groups |
1.60+0.08 |
|
Uronic acid |
4.05±0.20 |
|
Klason lignin |
30.34±3.32 |
|
Acid soluble lignin |
1.82±0.00 |
|
Ash |
1.41±0.16 |
|
Others (by difference) |
0.09 |
Table 4. Experimental design and results obtained by the hydrolysis of CAL of sunflower stalk by Trichoderma longibrachiatum xylanase
|
Run |
Variables |
Responses |
||
|
X1(mg ds/mL CAL) |
X2 (U/mL) |
Y1 (%) |
Y2 (g/g) |
|
|
1 |
50 |
150.00 |
57.37 |
9.64 |
|
2 |
50 |
150.00 |
57.15 |
9.66 |
|
3 |
75 |
60.00 |
40.78 |
7.42 |
|
4 |
25 |
240 |
58.20 |
8.18 |
|
5 |
50 |
277.26 |
55.51 |
9.15 |
|
6 |
50 |
150 |
59.09 |
9.46 |
|
7 |
50 |
22.74 |
37.42 |
8.71 |
|
8 |
50 |
150 |
53.74 |
9.62 |
|
9 |
25 |
60 |
51.39 |
9.90 |
|
10 |
14.65 |
150 |
58.56 |
8.78 |
|
11 |
75 |
240 |
57.75 |
9.11 |
|
12 |
50 |
150 |
55.16 |
9.47 |
|
13 |
85.35 |
150 |
50.13 |
7.67 |
|
X1: Substrat (g ds/mL CAL); X2: Enzyme(U/mL); Y1 (xylose yield)= 100 x (Xyl/Xylmax); Y2 (selectivity) = Xyl/Total other sugar relased from CAL during hydrolysis (Xyl/Glc). Xyl = xylose concentration obtained by the enzymatic hydrolysis of CAL, Xylmax= maximum xylose concentration in the CAL, Glc = glucose concentration obtained in the hydrolysate. |
||||
Table 5. Analysis of variance for xylose yield and selectivity
|
Source |
Sum of squares |
Degress of freedom |
Mean square |
F-value |
P-value |
|||||
|
Y1 |
Y2 |
Y1 |
Y2 |
Y1 |
Y2 |
Y1 |
Y2 |
Y1 |
Y2 |
|
|
Model |
544.42 |
7.47 |
5 |
5 |
108.88 |
1.49 |
33.00 |
102.36 |
0.0001 |
<0.0001 |
|
X1 |
66.02 |
1.22 |
1 |
1 |
66.02 |
1.22 |
20.01 |
83.35 |
0.0029 |
< 0.0001 |
|
X2 |
304.59 |
0.044 |
1 |
1 |
304.59 |
0.044 |
92.32 |
3.00 |
<0.0001 |
0.1267 |
|
X1 X2 |
25.81 |
2.91 |
1 |
1 |
25.81 |
2.91 |
7.82 |
199.16 |
0.0266 |
< 0.0001 |
|
X12 |
3.14 |
2.97 |
1 |
1 |
3.14 |
2.97 |
0.95 |
203.68 |
0.3618 |
< 0.0001 |
|
X22 |
147.97 |
0.63 |
1 |
1 |
147.97 |
0.63 |
44.85 |
43.24 |
0.0003 |
0.0003 |
|
Residual |
23.079 |
0.10 |
7 |
7 |
3.30 |
|
|
|
|
|
|
Lack of fit |
5.79 |
0.065 |
3 |
3 |
1.93 |
0.022 |
0.45 |
2.29 |
0.7332 |
0.2202 |
|
Pure error |
17.30 |
0.038 |
4 |
4 |
4.33 |
|
|
|
|
|
|
Total |
567.52 |
7.57 |
12 |
12 |
|
|
|
|
|
|
|
Xylose Yield |
R2 : 0.96, Adj R2 : 0.93, Pred R2 : 0.88, Adeq Precision : 16.7 C.V.% : 3.41, Std. Dev. : 1.82, PRESS : 68.2 |
|||||||||
|
Selectivity |
R2 : 0.99, Adj R2 : 0.98, Pred R2 : 0.93, Adeq Precision : 30.3 C.V.% : 1.35, Std. Dev. : 0.12, PRESS : 0.52 |
|||||||||
Table 6. The results for xylose yield and selectivity obtained at the optimum hydrolysis conditions and the predicted results for xylose yield and selectivity suggested by the program
|
Substrate (mg ds/mL CAL) |
Enzyme (U/mL) |
Yield (Predicted)(%) |
Selectivity (Predicted) (g/g) |
Yield (Actual) (%) |
Selectivity (Actual) (g/g) |
|
60 |
234 |
57.9 |
9.4 |
69.5+8.5 |
8.2+0.5 |
{kind=link}