Proteomics profiling has been approved as a powerful molecular strategy that has been widely implemented in dissecting the molecular basis of various biological processes in living organisms including plants. However, the experimental system and procedure of the employed proteomics profiling approach greatly affect the power and efficiency of proteomic profiling in dissecting the molecular mechanisms of a biological process [19-28]. In rice, seed development is a complex biological process that greatly affects grain yield and quality and is governed by complex regulatory networks comprising numerous transcription factors [29].
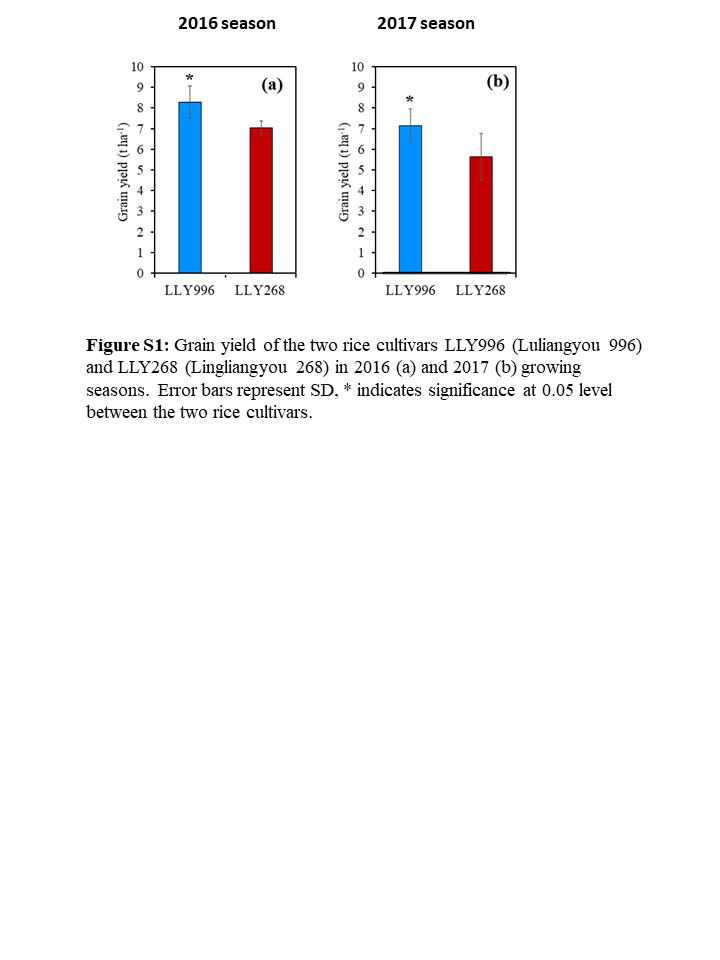
The current study has been carried out to uncover proteins implicated in amylose accumulation during the early period of grain filling in rice. Two rice cultivars, i.e., LLY996 than in LLY268, differed greatly in amylose accumulation during grain filling and in amylose content of mature grains were employed in the identification of differentially expressed proteins that might be implicated in amylose accumulation during grain filling. The data revealed that the cultivar LLY996 surpassed the cultivar LLY268 in grain-filling and amylose accumulation rates over the two growing seasons. Besides, amylose accumulation was significantly increased from the ninth day post-flowering until grain maturity in the cultivar LLY996 compared to the cultivar LLY268. These findings are consistent with previous results where a significant increment of amylose accumulation occurred primarily at 5–15 days post-flowering [30] and the highest rate of amylose accumulation occurred at 3–12 days post-flowering [31], indicating the suitability of grain samples collected 12 days post-flowering for the identification of proteins implicated in amylose accumulation during grain filling. Besides, the grain-filling and amylose accumulation processes were both well-fitted by the logistic equation for both cultivars, demonstrating the appropriateness of the two selected cultivars for quantitative proteomic profiling. The higher grain weight observed in the cultivar LLY996 compared to the cultivar LLY268 at maturity could be due to the higher-yielding ability of the cultivar LLY996 compared to the cultivar LLY268 (Figure S1) which is due to the genetic composition of the two cultivars (Figure 1).
In starch and sucrose metabolism, the enzyme isoamylase (ISA, EC: 3.2.1.68) is a starch debranching enzyme that has three isoforms, i.e., ISA1, ISA2 and ISA3, two of which, i.e., ISA1 and ISA3, are strongly implicated in amylopectin synthesis. Furthermore, although the ISA2 isoform appears to be catalytically inactive, it may modulate the action or stability of ISA1 [32]. However, all three isoforms reduce granular starch, where amylose synthesis occurs within the granules [32, 33]. In our study, the differentially expressed protein LOC_Os05g32710.1 (ISA2) was down-regulated in the cultivar LLY996 and up-regulated in the cultivar LLY268 (Table 4), which is similar to a previous report of ISA as a starch debranching enzyme that has been up-regulated in a low-amylose content rice mutant [14]. The accumulation rate of amylose is positively correlated with the activity of the Granule-bound starch synthase (GBSS) enzyme [34]. A similar relationship was observed in our study where the protein associated with LOC_Os06g04200.1 which is involved in GBSS activity has been up-regulated (3.39-fold change) in the cultivar LLY996 compared to the cultivar LLY268 (Table 4). These findings suggest the implication of the locus LOC_Os06g04200.1 in enhancing amylose synthesis and accumulation in rice.
Several differentially abundant proteins have been identified to be implicated in the glycolysis and gluconeogenesis which involve reversed biochemical reactions of each other’s pathways and most of the associated enzymes take part in reversible reactions of the pathways [14, 35]. Glucose-6-phosphate isomerase (EC: 5.3.1.9) catalyzes the glucose-6-phosphate and fructose-6-phosphate and the reaction is reversible [36]. Triosephosphate isomerase (EC: 5.3.1.1) is involved in sugar metabolism and, basically, the pathway of glycolytic synthesis of ATP [37]. The aldose-1- epimerase protein is the key enzyme (EC: 5.1.3.3) of carbohydrate metabolism and catalyzes the interconversion of α- and β-anomers of sugar [38]. The key enzyme 6-phosphofructokinase, which is a pyruvate kinase and pyruvate phosphate dikinase catalyze irreversible reactions in glycolysis [14], did not show differential expression between the two cultivars, suggesting that there was no Gluconeogenesis involved during grain filling and amylose accumulation. There was three differentially expressed proteins, i.e., LOC_Os01g62420.1, LOC_Os02g36600.1, and LOC_Os08g37380.2, which exhibited 0.826, 0.725, and 0.662 fold changes in the cultivar LLY996 compared to the cultivar LLY268 and are known to be implicated in triosephosphate isomerase, aldose-1- epimerase, and glucose-6-phosphate isomerase, respectively (Table 4). These three enzymes limit the glycolytic process and decreased the glucose-1-phosphate consumption.
Glucose-1-phosphate is a key factor that links glycolysis/gluconeogenesis and starch and sucrose metabolism (Figure 4). Glucose-1-phosphate is one of the first substrates in starch synthesis and glycolysis [39, 40]. It is one of the substrates that are catalyzed to form adenosine diphosphate glucose (ADPG), then the ADPG is catalyzed by GBSS to elongate amylose [13, 14]. In our study, we hypothesized that uridine diphosphate glucose (UDPG) obtained single glucose from glucose-1-phosphate and then was catalyzed by GBSS to form amylose. Reportedly, UDPG could be converted into hexose phosphates and take on roles in starch synthesis [40, 41]. In the process of amylose synthesis, maltohexose acts as one form of primers in plants [16]. However, what still remains to be investigated is whether UDPG provides the hexose or, similar to the role of ADPG, provides the single glucose molecule for the primer to elongate amylose. The use ratio of glucose-1-phosphate in starch and sucrose metabolism and glycolysis also has to be further studied.
Glycolysis and gluconeogenesis involve nearly reversed biochemical reactions of each other’s pathways [35] and most of the associated enzymes take part in reversible reactions of the pathways [14]. Glucose-6-phosphate isomerase (EC: 5.3.1.9) catalyzes the glucose-6-phosphate and fructose-6-phosphate and the reaction is reversible [36]. Triosephosphate isomerase (EC: 5.3.1.1) is involved in sugar metabolism and, basically, the pathway of glycolytic synthesis of ATP [37]. The aldose-1- epimerase protein is the key enzyme (EC: 5.1.3.3) of carbohydrate metabolism and catalyzes the interconversion of α- and β-anomers of sugar [38]. The key enzymes 6- phosphofructokinase, pyruvate kinase and pyruvate phosphate dikinase catalyze irreversible reactions in glycolysis [14] and we did not observe differences in the expression of any of these enzyme between the two cultivars, which suggests there was no Gluconeogenesis occurring.
{kind=link}