This pilot study represents one of the first unbiased proteomic analyses on pre- and postoperative CSF samples from patients with perioperative neurocognitive disorders compared to controls. This approach offers a powerful tool for characterizing CSF protein content in the perioperative setting and has the potential to identify pathways involved in the mechanisms underlying POCD, delirium and other perioperative neurocognitive disorders. We identified peptides from 11 proteins with statistically significant intensity changes (q < 0.05) in our FDR-adjusted linear mixed model, and pathway analysis revealed trend-level differences in 50 peptides from 17 complement and coagulation-related factors, suggesting a possible role for these pathways in POCD pathogenesis. Since we planned this study to evaluate the CSF proteome without focusing specifically on either these 11 proteins or the complement pathway, our findings illustrate the strength of unbiased CSF proteomic analyses for identifying potential “unknown unknowns” in the pathogenesis of perioperative neurocognitive disorders. Additionally, our findings on the variability of CSF peptide levels in older surgical patients may prove useful for guiding power calculations when planning future studies.
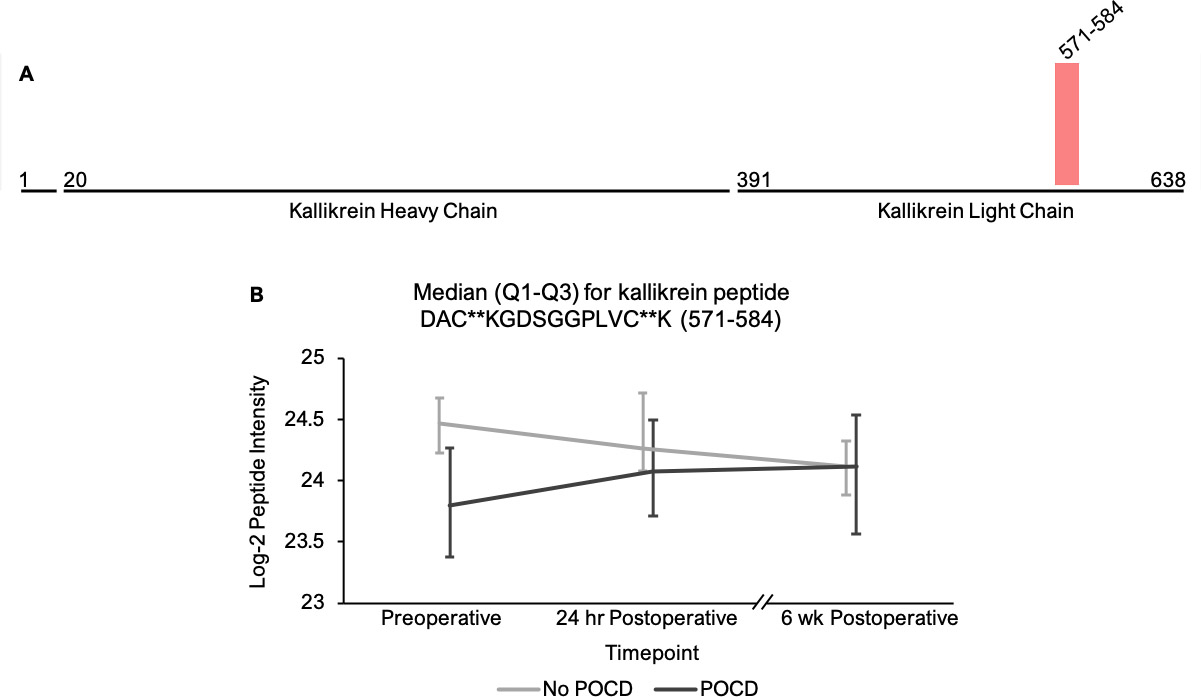
We identified 12 peptides from 11 proteins that satisfied the stringent significance threshold of q < 0.05 (Supplemental Table 1) in our linear mixed model examining an interaction between groups (POCD vs no POCD) and postoperative time points. Several of these 11 proteins have been previously implicated in other neurocognitive disorders. First, cystatin C appears to play a neuroprotective role in Alzheimer’s disease, possibly through inhibition of cerebral amyloidosis[51, 52]. Furthermore, low cystatin C levels and certain cystatin C polymorphisms have been identified as predisposing factors to Alzheimer’s disease (reviewed)[53] and contribute to poor cognitive outcomes in Parkinson’s disease[54], after ischemic strokes[55], and in HIV-associated neurocognitive disorder[56]. Similarly, multiple peptides representing the copper and iron-handling protein ceruloplasmin were found to vary significantly between groups at q < 0.05. Copper storage and regulation abnormalities have been implicated in the pathogenesis of Alzheimer’s disease (reviewed)[57] and HIV-associated neurocognitive disorder[58], and alterations in ceruloplasmin activity have been associated with increased Alzheimer’s disease risk[59, 60]. Finally, the axonal secretory sorting receptor secretogranin-III, represented by a single peptide with q < 0.05, has been implicated in the mechanisms driving amyloid-mediated neurodegeneration in Alzheimer’s disease[61]. To our knowledge, these proteins have not previously been previously associated with POCD. Targeted proteomic studies to determine their roles in perioperative neurocognitive disorders are thus warranted.
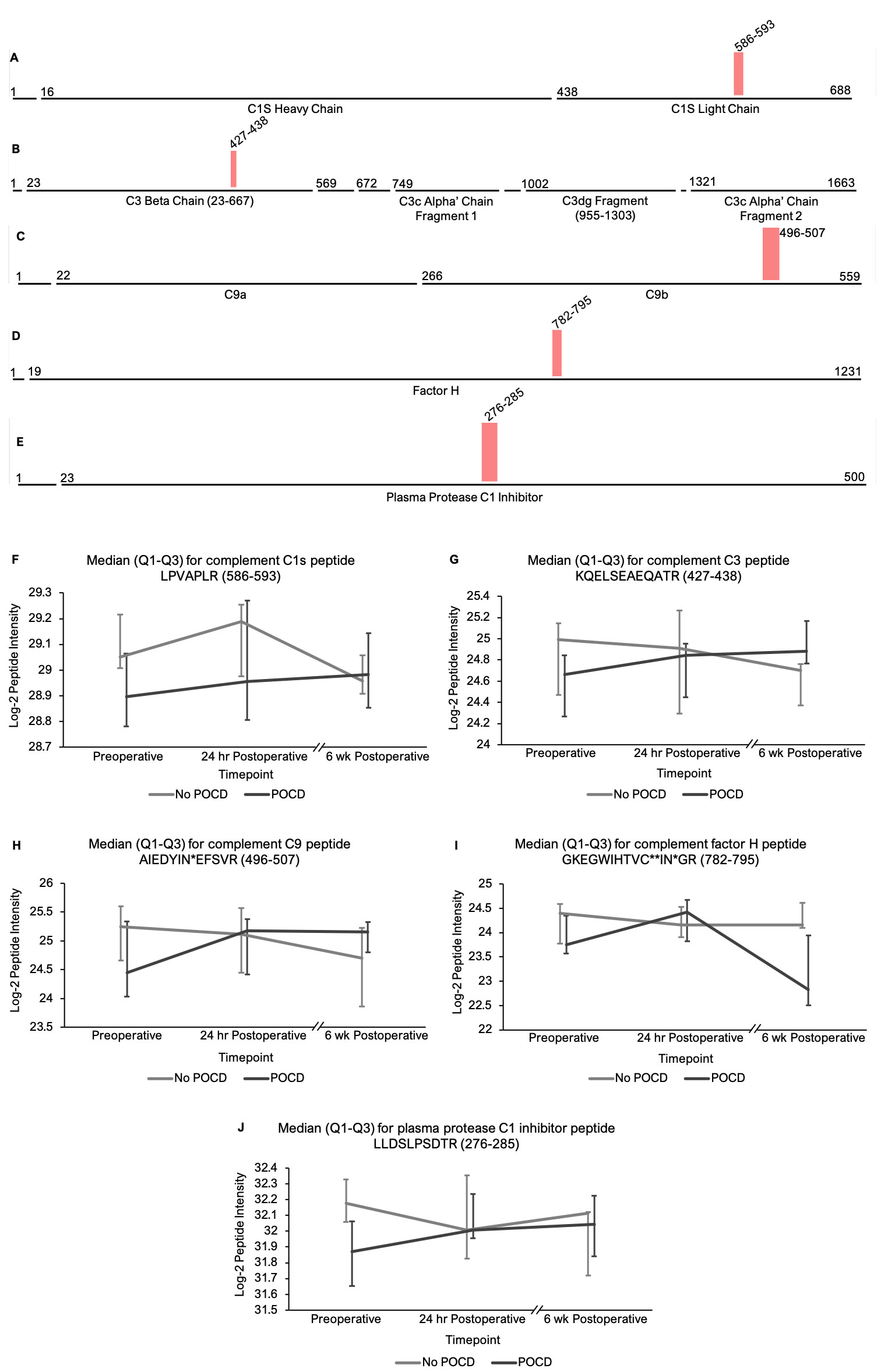
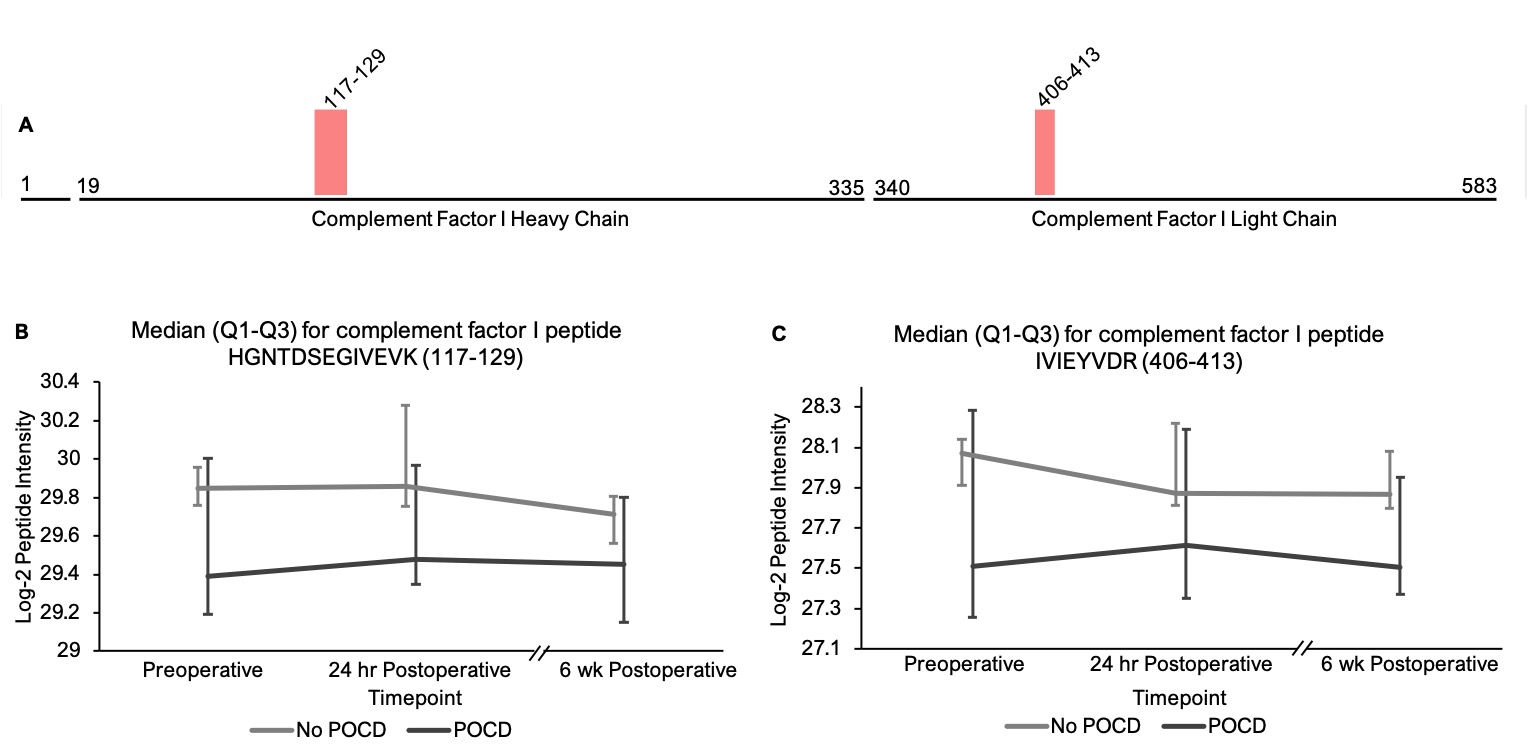
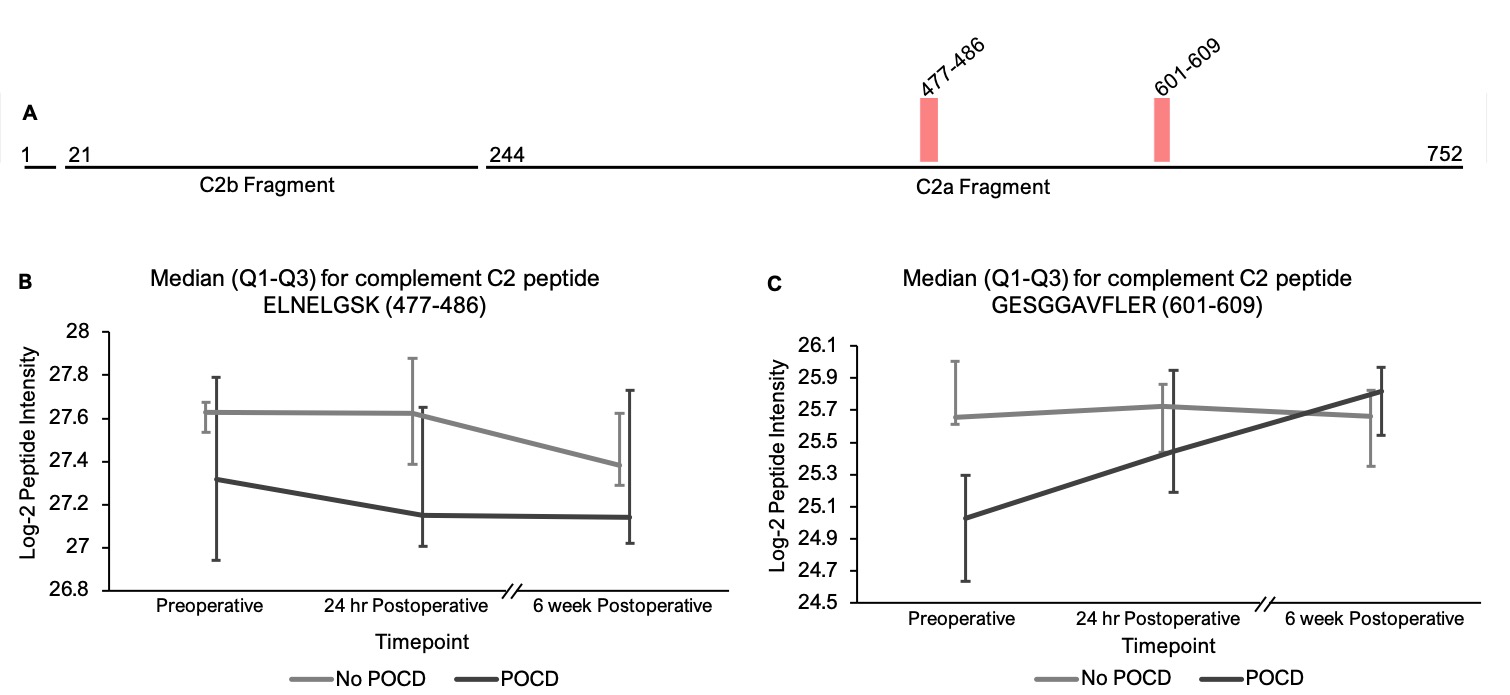
Pathway analysis on peptides that showed trend-level significance with q < 0.25 identified a significant role for the complement and coagulations factors pathway(s). The hypothesis that complement plays a role in perioperative neurocognitive disorders aligns with current animal models of surgery-induced neuroinflammation and cognitive decline[62]. Complement regulates brain development and homeostasis by driving microglial synaptic phagocytosis[63], and in murine models, complement-dependent synaptic pruning is critical for optimizing cognitive performance by reducing excessive synapse numbers that can cause epilepsy[64, 65]. However, abnormal CNS complement activity has also been revealed in several neurocognitive disease states as a neuroinflammatory trigger for microglial activation and neuronal damage[66–68]. A role for complement in perioperative neurocognitive disorders would fit the theory that these disorders are driven by neuroinflammation[1, 69]. Indeed, murine model studies have shown that complement C3 levels, an initiator of the complement cascade, and levels of the receptor for its cleavage product (C3aR) are elevated after orthopedic surgery, and these increases have been shown to contribute to cognitive dysfunction in mouse POCD models[62]. Similarly, elevated baseline CSF C3 levels have been associated with higher rates of postoperative delirium in human patients[70], and critically-ill patients with delirium have been found to exhibit upregulated CSF complement levels[71]. Our results align with these prior studies to support a role for dysregulated complement in human perioperative neurocognitive disorders.
Complement pathway activation has been observed in peripheral blood after cardiac surgery involving cardiopulmonary bypass[72], which prompted a study on the effect of the anti-C5 monoclonal antibody pexelizumab on POCD rates following coronary artery bypass graft surgery[73]. This study measured changes in cognitive function from before surgery to 4 and 30 days after surgery, and found reduced visuospatial cognitive deficits at both time points in patients who received pexelizumab compared to controls[73]. Pexelizumab had no effect on global cognition at either timepoint, though this negative finding is unsurprising for two reasons: first, it was administered only for 24 hours and has a relatively short half-life (20–27 hours)[74]. Second, the penetration of monoclonal antibodies across the blood-brain barrier is limited[75], and it is unknown whether the mild increase in blood-brain barrier permeability due to anesthesia[76] would permit sufficient CNS access for pexelizumab to effectively block complement activity within the brain. Thus, a proper test of the role of complement activation in POCD will require future studies utilizing complement inhibitors that cross the blood-brain barrier and which have sufficient half-lives.
Our ability to identify additional proteins and pathways associated with POCD in this pilot study was limited by a high degree of variance in peptide intensities among the participants, and there was no peptide that showed intensity differences between groups at any single time point. Despite this limitation, analysis of our results revealed two noteworthy trends. First, among the all complement proteins containing peptides with significantly different intensities in our mixed models, the mean baseline peptide levels were lower in patients who later developed POCD. Second, over the course of six weeks, there was a clear trend toward convergence of these peptide intensities for all of the proteins except complement factor H, which further diverged by the six-week time point. The consistency of these trends, despite high peptide intensity variance within groups, suggests that significant results will likely be detected in a larger cohort. Thus, the data collected in this study may prove useful for planning future studies using unbiased proteomic analyses on perioperative CSF samples. For example, a power analysis on complement C5 and C6 peptide intensities calculating for α = 0.0001 to account for multiple testing indicates that group sizes of > 31 patients would have 80% power to reveal significant differences between baseline peptide intensities (calculated for C6 peptide KALQEYAAK with a difference of 1.419*107 between means). Group sizes of 108 would have at least 80% power to detect baseline intensity differences in half of the C5 and C6 peptides with q < 0.25 in our linear mixed model.
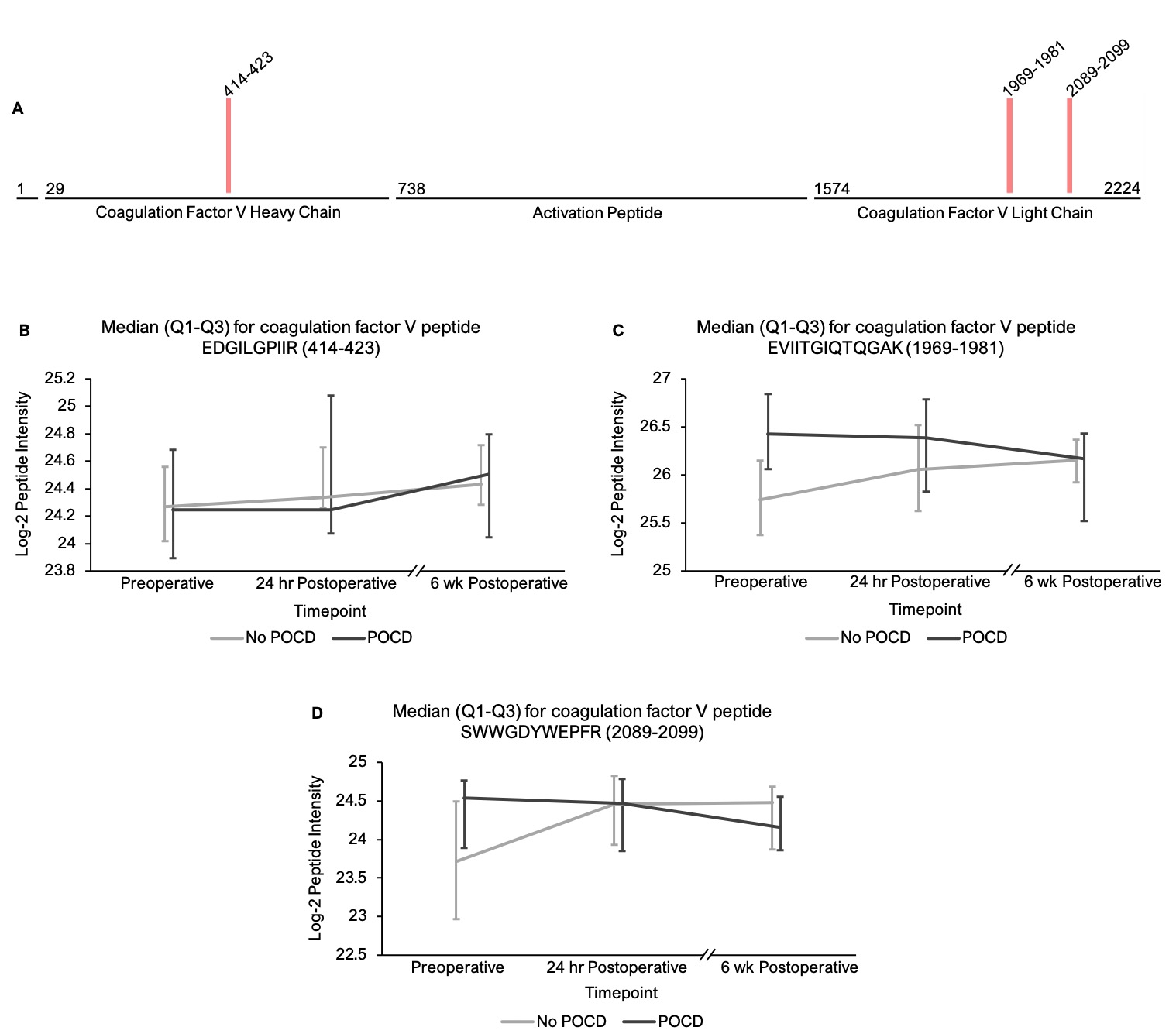
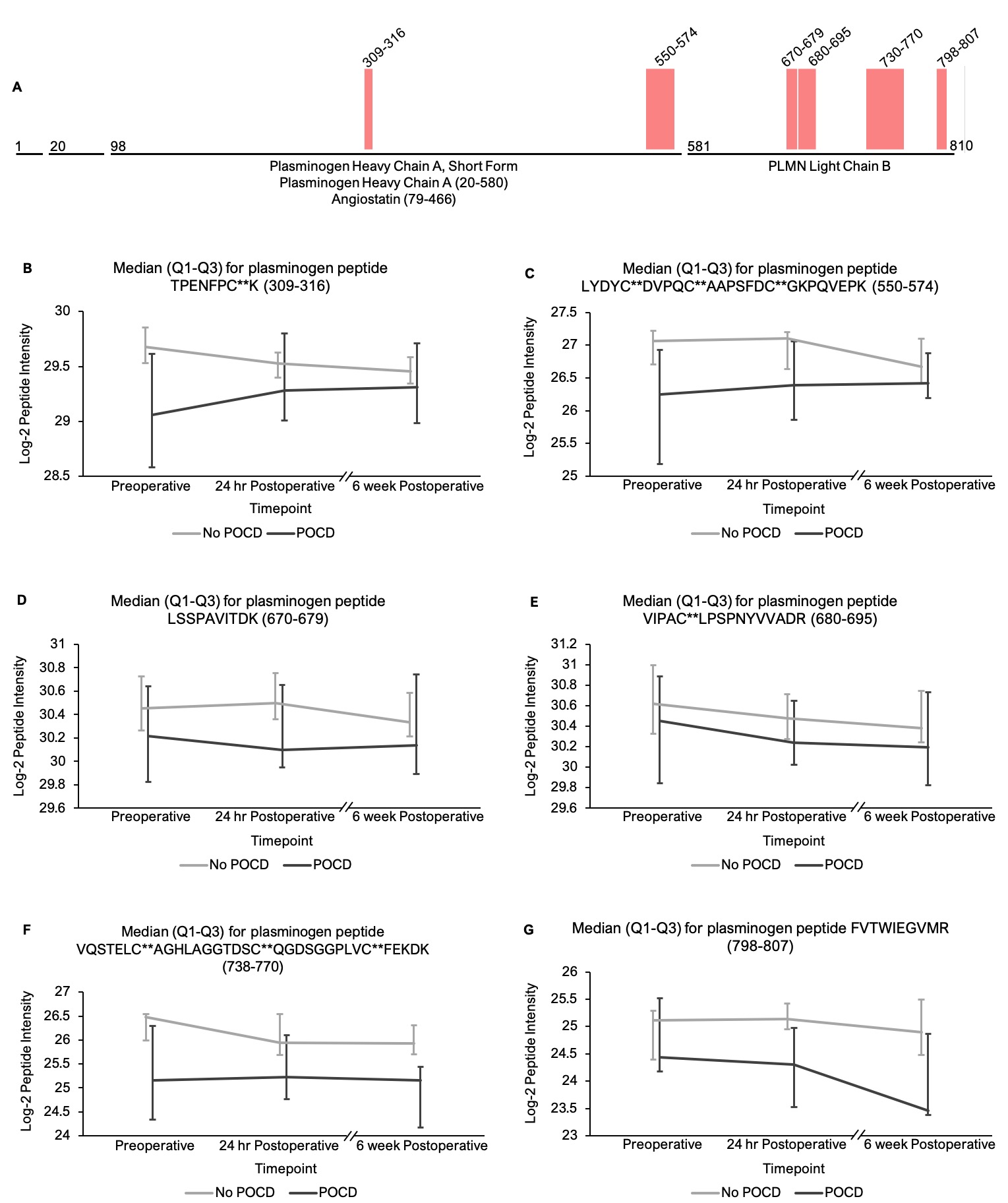
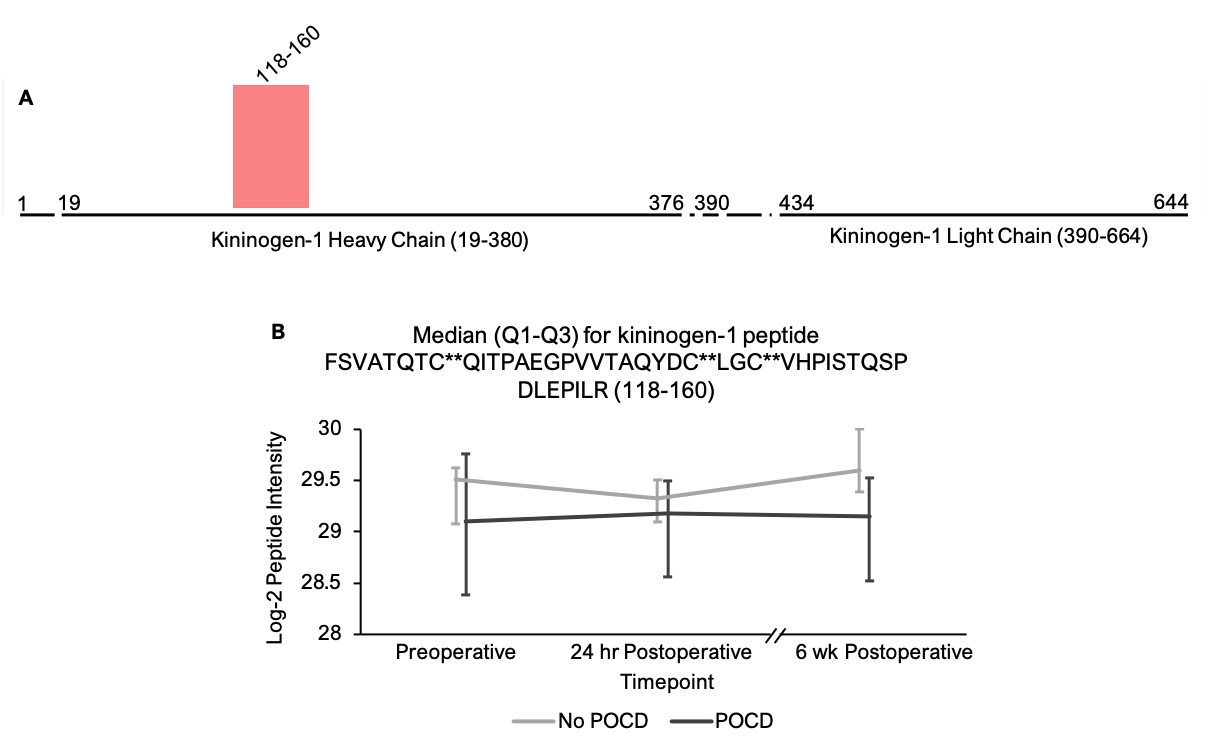
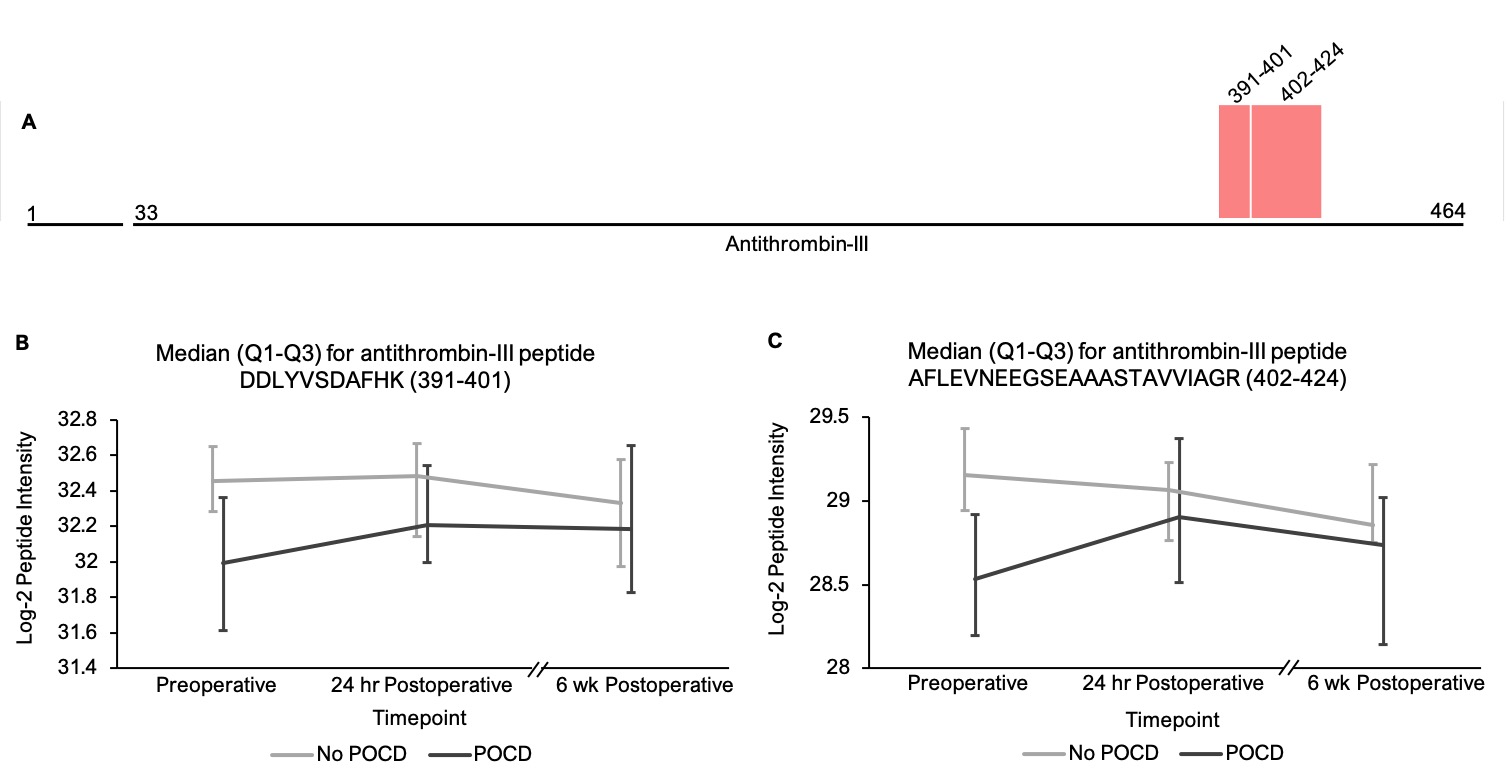
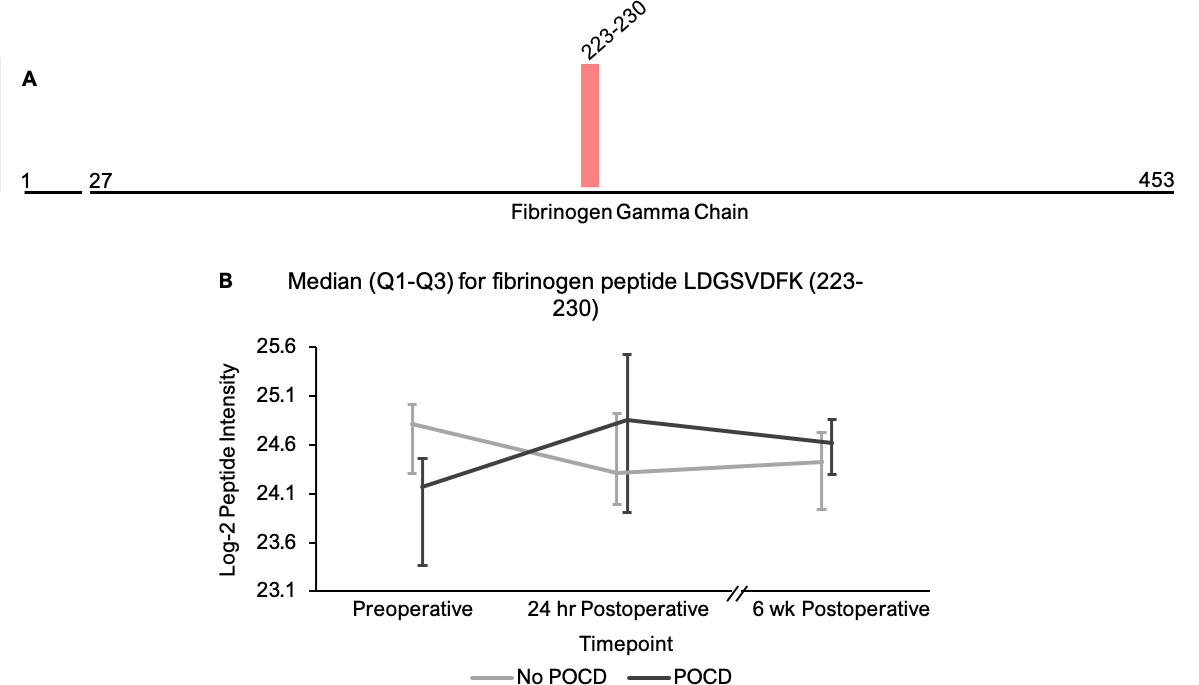
In addition to the complement peptides that differed between groups, several non-complement factors with known pro-inflammatory and/or hemostatic functions were identified by our linear mixed models as trending toward significance (q < 0.25) between the patients with versus without POCD over the six-week period. Notably, fibrinogen and its cleavage products are known to contribute to inflammation in the CNS by a variety of mechanisms including microglial activation[77, 78], neuronal damage[79], interactions with beta-amyloid[80], and disruption of the blood-brain barrier[81]. In fact, alterations in CNS fibrinogen biology have been implicated in the pathogenesis of neurological, neurocognitive, and psychiatric disorders including multiple sclerosis[82], traumatic brain injury[83], Alzheimer’s disease[78, 84], and depression[85]. Likewise, coagulation factor V, a central mediator of hemostasis, has been previously linked to delirium pathogenesis[33]. Further, plasminogen has been shown to potentiate neuroinflammation[86, 87] and Alzheimer’s disease-related pathology[86], and elevated serum kininogen has been associated with depression[85]. As with complement, our data suggest a relationship between POCD and dysregulation of these inflammatory and thromboactive factors.
Several limitations apply to this study. First, although the CSF analyses presented here suggest differential changes in peptides levels from complement and coagulation-related proteins between patients with versus without POCD, it is possible that these changes could reflect non-causal associations. In essence, these pathways may be markers of POCD without playing an active role in causing it. Second, like other mass spectrometry-based analyses, the intensities measured in this study reflect relative peptide levels rather than quantitative concentration measurements. Thus, while our unbiased approach has identified candidate proteins with suspected involvement in POCD, precise protein levels and inter-protein comparisons will require additional targeted studies (i.e. proteomic studies with internal controls designed for precise quantification). Third, the results of this preliminary study are by no means definitive, as larger future studies are needed to more thoroughly assess the role of these proteins and pathways in POCD and/or other perioperative neurocognitive disorders. Fourth, it is unclear whether the differential changes in CSF protein levels observed here are driven primarily by changes in expression, activity, or turnover initiated within the CNS, versus peripheral inflammation and activation of the coagulation and complement cascades after surgery that crosses the blood-brain barrier into the CNS. Further research comparing postoperative changes in both the CSF and plasma proteomes at multiple simultaneous time points may help evaluate these two different possibilities.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}