Rice CGTs responsible for varietal di- C -glycosides
In our ongoing investigation of Gramineae CGTs, we first compared the differences of C-glycoside spectrum between two rice subspecies (japonica vs indica) in detail (Fig. 1a). The rice leaves were extracted and subjected to LC-MS/MS analysis. Because most of the flavone C-glycosides in rice share the common aglycone apigenin (Api) or luteolin (Lut) (Besson et al. 1985), we determined to focus on five representative groups of Api/Lut-C-glycosides, corresponding to monopentosides, monohexosides, dipentosides, pentosylhexosides and dihexosides. Both rice varieties were found to predominantly produce di-C-glycosides (96% in O. sativa japonica, 91% in O. sativa indica, Fig. 1a), however, the composition of diglycosides differed drastically. The japonica rice particularly accumulated apigenin C-pentosylhexoside (corresponding to m/z [M–H]− = 563.1), whereas the indica rice majorly produced apigenin di-C-pentoside (corresponding to m/z [M–H]− = 533.1) besides apigenin C-pentosylhexoside (Fig. 1a). The most abundant diglycosides were verified as schaftoside (Sch) and apigenin 6,8-di-C-arabinoside (Api-di-C-Ara) as referenced to the authentic samples (Fig. 1b). In accordance with the previously recorded metabolic profiling (Kim et al. 2018a; Narasimhan et al. 1989; Yang et al. 2016), glucosyl and arabinosyl residues seem to be the representative hexose and pentose present in rice. Other minor flavone diglycosides were proposed to be C-pentosylhexosides, di-C-pentosides and di-O-glycosides with diverse sugar-linkages (Fig. S2). It is also worthy to note that C-glycosides of apigenin are generally more abundant than those of luteolin, regardless of the glycosylation patterns (Fig. 1a).
In our previous work, both Clade A and Clade B CGTs from grass family were proved to be able to recognize the non-sugar-bearing aglycones [i. e., phloretin, 2-hydroxynaringenin (2-OHNar) (Sun et al. 2020), resulting in majorly mono-C-glucosides and arabinosides. It remains unclear that in rice how the aglycones undergo two steps of C-glycosylation to reach di-C-glycosides bearing different sugars (for example, schaftoside and isoschaftoside). According to the existing knowledge of flavone C-glycoside biosynthesis (Putkaradze et al. 2020), we proposed a biosynthetic pathway in which the rice CGTs collaborate to first install a C-glucosyl (mainly by Clade A CGTs) on the precursor (2-OHNar), followed by addition of a second C-arabinosyl group (mainly by Clade B CGTs) (Fig. 2, black arrows). To verify whether Clade B CGTs cloud accept monoglucoside substrates like C-glucosyl-2-hydroxynaringenin (C-Glc-2-OHNar), we expressed the His6-tagged OsUGT708A1, OsUGT708A2, OsUGT708A39, and OsUGT708A40 in E. coli BL21(DE3) and tested their activities toward nothofagin (3’-C-glucosyl phloretin, a relatively stable analogue of C-Glc-2-OHNar) in in vitro enzymatic assays. At the presence of UDP-Ara, nothofagin was converted to a new product with m/z = 567.2 (Fig. 1c, 1d). The characteristic fragment ions such as [M–H–150]−/[M–H–180]−/[M–H–210]− in MS/MS spectrum clearly revealed a hybrid pattern of C-pentosylation and C-hexosylation in good agreement with the structure of 3’-C-glucosyl-5’-C-arabinosyl phloretin. These results suggested that rice Clade B CGTs could catalyze the arabinosyltransferring reaction of C-monoglucoside substrate, which was a key step in the biosynthesis of C-pentosylhexosides like schaftoside or isoschaftoside. Recently, the dissection of schaftoside pathway in other plants (mainly represented by dicot plants like Scutellaria baicalensis and Nelumbo nucifera) also supported the above results (Feng et al. 2021; Wang et al. 2020), which shows the generality of di-C-glycoside pathway in higher plants.
Unlike O. sativa japonica that accumulates C-pentosylhexoside, O. sativa indica produces a large amount of apigenin di-C-pentoside occupying 46% of the total flavone C-glycosides (Fig. 1a). There is also an increase of apigenin mono-C-pentoside (corresponding to m/z [M–H]− = 401.1). This is probably due to the three additional clade B CGTs (OsUGT708A1, OsUGT708A39 and OsUGT708A40) only present in indica rice (Fig. 1), which can utilize UDP-Ara to convert 2-OHNar to C-Ara-2-OHNar (alternatively, phloretin to C-arabinosyl phloretin) (Sun et al. 2020). In particular, among the Clade B CGTs, OsUGT708A40 is a unique di-C-arabinosyltransferase that catalyzes a tandem C-arabinosylation reaction (Sun et al. 2020). We proposed that OsUGT708A40 was a key di-C-arbinosyltransferase responsible for the formation of apigenin 6,8-di-C-arabinoside (Fig. 2, light blue arrows)
Introduction of UDP-arabinose and UDP-xylose supply allowed de novo biosynthesis of schaftoside, isoschaftoside, vicenin-1 and vicenin-3
To further prove our proposed pathway (Fig. 2a, 2b) and achieve de novo biosynthesis of bioactive di-C-glycosides, we selected the fast growing and genetically amenable E. coli as a suitable chasis for pathway reconstitution. The previously constructed sCZ112 harboring pYH55 (Li et al. 2019) and pCZ201 (Sun et al. 2020) for optimized 2-hydroxynaringenin production was used as the starting strain. In order to realize the heterologous biosynthesis of C-pentosylhexoside like schaftoside, we first assembled a di-CGT cassette containing PhUGT708A43 (an excellent coding C-monoglucosylating enzyme from moso bamboo (Sun et al. 2020) for the first step of glucosylation) and OsUGT708A1 (for the subsequent C-arabinosylation) under T7 promoter (Fig. 3a).
A major difficulty for the biosynthesis of arabinosides in E. coli is the absence of native UDP-arabinose supply. To solve this problem, we introduced SmUxs (UDP-xylose synthase) and SmUxe (UDP-xylose 4-epimerase) from Sinorhizobium meliloti 1021 (Gu et al. 2011) to enable the metabolism from UDP-glucose to UDP-arabinose (Fig. 2a). Two SmUxs homologues (SmUxs1 and SmUxs2), sharing only 57.3% amino acid identity, were respectively ligated downstream to the PhUGT708A43-OsUGT708A1 cassette and further assembled with SmUxe to give pCZ193-1 and pCZ193-2 ready for the production of schaftoside (Fig. 3a). After transferring pCZ193-1 or pCZ193-2 into sCZ112 (resulting in strain sCZ113 and sCZ114, respectively), we successfully detected 2.75 mg/L schaftoside (Sch) and 0.43 mg/L isoschaftoside (Isosch) in sCZ113 broth through 72 hours fermentation in MOPS media (Fig. 3b). The pathway intermediates like vitexin (Vit, 15.14 mg/L), isovitexin (Isovit, 9.78 mg/L), naringenin (Nar, 45.54 mg/L) and p-coumaric acid (p-CA, 34.79 mg/L) were also observed (Fig. 3a, 3b). All the products were identified through comparison with authentic samples in HPLC analysis (Fig. 3b) and high-resolution (HR) MS/MS spectroscopic data (Fig. 3c, Fig. S3). On the other hand, 2.67 mg/L Sch and 0.41 mg/L Isosch were detected in sCZ114. The accumulation of Vit, Isovit and Nar reached 14.52 mg/L, 10.42 mg/L and 38.01 mg/L. A similar productivity of Sch/Isosch and no significant difference of accumulation pattern of intermediates between SmUxs1 and SmUxs2 (Fig. 3a), therefore we used SmUxs1 for further experiments.
Since UDP-xylose is an upstream precursor of UDP-arabinose (Fig. 2a), we proposed that flavone C-xylosides might be generated in a truncated pathway containing biosynthetic genes fitting just for UDP-xylose biosynthesis (Fig. S4). Therefore, we also try to achieve the production of vicenin-1 (apigenin 6-C-xylosyl-8-C-glucoside, Vic-1) and vicenin-3 (apigenin 6-C-glucosyl-8-C-xyloside, Vic-3). After transferring pCZ192-1 (harbors the cassette of PhUGT708A43-OsUGT708A1-SmUxs1) into sCZ112 (resulting in strain sCZ115), we detected a trace amount of Vic-1 (0.09 mg/L) and Vic-3 (0.28 mg/L) in 72 hours fermentation (Fig. S5), which is a much lower titer compared to that of Sch and Isosch. This result indicated that UDP-xylose might not be a favorite sugar donor of OsUGT708A1. To the best of our knowledge, di-C-glycosides like Sch, Isosch, Vic-1 and Vic-3 were synthesized in heterologous chassis cells for the first time. These results indicated the feasibility of de novo production of C-arabinoside and C-xyloside in E. coli.
De novo biosynthesis of apigenin di-C-arabinoside and minor C-pentosides
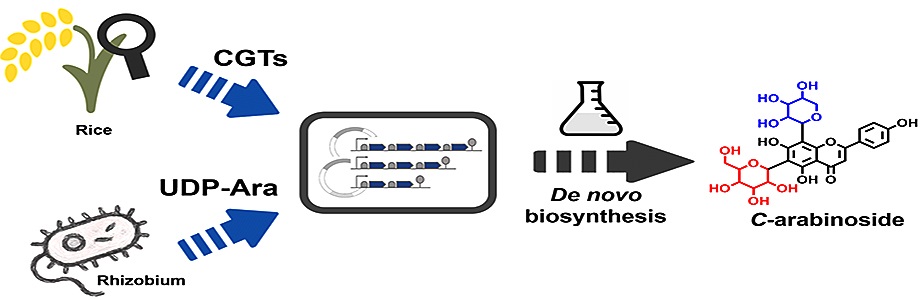
Flavone compounds bearing multiple C-pentosyl (for example, arabinosyl, xylosyl) residues are uncommon natural products. To further expand the diversity of flavone C-glycosides, we attempted to construct an artificial pathway in E. coli for the production of apigenin di-C-arabinoside and other minor C-pentosides. The biosynthesis of specific di-C-arabinosides requires efficient di-C-glycosyltransferase preferring UDP-Ara, as well as a heterologous UDP-Ara-synthesizing module above-mentioned. OsUGT708A40 was selected as a proper enzymatic part since it was identified as the only di-C-arabinosyltransferase in rice (Sun et al. 2020). Due to the close similarity of UDP-Ara and UDP-xylose (UDP-Xyl), we predicted that OsUGT708A40 might also promiscuously consume UDP-Xyl for some minor C-xyloside production.
The construct pCZ194 harboring OsUGT708A40-SmUxs1-SmUxe cassette were transformed into sCZ112 to give strain sCZ118 (Fig. 4a). After 72 hours fermentation, we detected the emergence of new products characteristic of C-pentosides instead of C-glucosides (i.e., Vit/Isovit) in the fermentation media. We quantified these products using vitexin and vicenin-2 as internal standards. The major peak with a retention time (Rt) = 16.02 min was identified as apigenin 6,8-C-di-arabinoside (Api-di-C-Ara) based on both LC-MS/MS and NMR evidence (Fig. 4b, Fig. S6). The 1H NMR spectrum of Api-di-C-Ara recorded at 80 ℃ clearly revealed the doublets of aglycone H2’, 6’ and two anomeric protons of sugars (Fig. S7). The α-l-arabinosyl was clarified though correlations between Ara-H1, Ara-H5a and Ara-H4 observed in 2D NOESY spectrum (Fig. 6f). In addition to Api-di-C-Ara, we also detected two minor C-pentosides which are supposed to be apigenin 6,8-C-di-xyloside (Api-di-C-Xyl) and chrysin 6,8-C-di-arabinoside (Chr-di-C-Ara), according to the HR-MS/MS and NMR analyses (Fig. 4c, Fig. S8). Unfortunately, we didn't succeed in obtaining and distinguishing apigenin 6-C-arabinoside or 8-C-arabinoside [Api-(6/8)-C-Ara], since they decomposed quickly in the solvent (Fig. S9). Without extra optimization, the titer of Api-di-C-Ara, Api-di-C-Xyl, Chr-di-C-Ara and nascent Api-C-Ara reached 24.89 mg/L, 0.78 mg/L, 0.38 mg/L, 21.15 mg/L, respectively.
Similarly, we also constructed pCZ195 specific for Api-di-C-Xyl production (Fig. S8). As expected, after 72 hours fermentation, we detected 3.26 mg/L Api-di-C-Xyl as major product with no flavone C-arabinosides accumulated (Fig. S10). While compared with the productivity of Api-di-C-Ara (24.89 mg/L) in sCZ118, the production of Api-di-C-Xyl was much lower. This could also be explained by the substrate preference of OsUGT708A40 to UDP-Ara rather than to UDP-Xyl.
Fed-batch fermentation of C-arabinosides
To achieve a large-scale production and verify the scalability of our C-glycoside-producing strains, we performed scale-up fermentation of sCZ113 and sCZ118 in a 5 L bioreactor. The minimal M9 media with 20 g/L glucose was used as basal culture medium and 500 g/L glucose was used as supplementary medium. During the fermentation process of sCZ113, p-coumaric acid (p-CA) rapidly accumulated to 66.1 mg/L at 9 hours (after induction) at the first stage and then rapidly decreased (Fig. 5a). Afterwards, naringenin (Nar) accumulated to 75.8 mg/L at 16 hours until it was consumed. Vitexin (Vit) and isovitexin (Isovit) appeared at about 9 hours, and schaftoside (Sch)/isoschaftoside (Isosch) appeared later (at approximate 16 hours). After 81 hours fermentation, production of Sch and Isosch reached 19.87 mg/L (7.2 folds compared to flask-shake) and 2.41 mg/L (5.6 folds compared to flask-shake) with 22.87 mg/L Vit and 13.32 mg/L Isovit left.
During the fermentation process of sCZ118, p-CA (74.8 mg/L) and Nar (20.29 mg/L) first rapidly accumulated to the maximum within 9 hours (Fig. 5b). After 84 hours fermentation, production of Api-di-C-Ara reached to 113.16 mg/L (4.7 folds compared to flask-shake). The results confirm that our fermentation process could be scaled up controllably and productively, which proved that fed-batch fermentation was beneficial to the accumulation of downstream glycosylated products. Our engineered E. coli system possess the ability to supply enough UDP-Ara for large production of flavone C-arabinosides, which displays great industrial potential.
{kind=link}