The human Y chromosome is a male sex-determination chromosome and escapes meiotic recombination. Despite its small size and limited gene content, the Y chromosome has crucial roles in development, differentiation, as well as gender-based diseases, which are often confined to the hypothesis (Bellott et al., 2014).
The Y chromosome harbors 47 protein-coding genes, where 35 of them have protein evidence (PE) at protein level (PE1), 5 proteins at the transcript level (PE2), and 7 genes encoding uncertain proteins (PE5) (https://www.nextprot.org/about/protein-existence). Therefore, Chromosome-centric Human Proteome Project (C-HPP) was launched in 2012, to identify human missing proteins and investigate their functions. (Meyfour et al., 2019). The human Y Chromosome Proteome Project (Y-HPP) is trying to recognize the functions of the Y chromosome’s proteins (Jangravi et al., 2013).
In pursuit of this goal, several studies have been undertaken, including a tumor suppressor role for KDM5D in the prostate cancer cell line (Jangravi et al., 2015) and new roles for DDX3Y in neural differentiation of NTERA-2 human embryonal carcinoma cells (NT2) (Vakilian et al., 2015), as well as TBL1Y and also KDM5D in cardiac differentiation of hESC (Meyfour et al., 2019, Meyfour et al., 2017a).
Since many genes are silent in adult cells, investigating the functions of their proteins is challenging. To meet this challenge (pluripotent stem cells) PSCs might be used as a valuable tool to achieve proteins and missing proteins function, which is also a target of the Y chromosome project (Alikhani et al., 2020).
These studies highlighted the remarkable role of some genes that help to candidate the target genes for more investigations. Among genes of interest, theexpression of RPS4Y1 showed sexual dimorphism of the kidney, and SRY mediated kidney development and played a role in the blood pressure control in males (Meyfour et al., 2017b).
The expression of RBMY1, HSFY1, RPS4Y1, and SRY increased during the neural cell differentiation of the NT2 cell line (Vakilian et al., 2015). Furthermore, The overexpression of RBMY1, HSFY1, RPS4Y1, and several other MSY genes, was observed at the transcriptional level during the cardiac differentiation of hESCs (Meyfour et al., 2017a).
Genes located in the azoospermia factor (AZF) regions have a critical role in spermatogenesis and fertility, and AZF microdeletion can impair their functions. Our target genes, RBMY1A1 and HSFY1, are in the AZF region. The RBMY1 family consists of approximately 30 genes and pseudogenes in six subgroups on the Y chromosome. RBMY1A1 contains a conserved domain called the SRGY motif (serine, arginine, glycine, and tyrosine) that modulated its function. RBMY1 plays a significant role in cardiac development, spermatogenesis, infertility, prostate cancer, and hepatocellular carcinoma (HCC) (Meyfour et al., 2017b, Chua et al., 2015, Kido et al., 2020, Jangravi et al., 2013, Dasari et al., 2001).
The Heat Shock Factors (HSFs) are a family of transcription factors that encode chaperones (Duchateau et al., 2020). Their critical role is stress responses in abnormal conditions such as oxidative stress, thermal stress, hypoxia, and protein degradation (Chatterjee and Burns, 2017). They also participate in gametogenesis, embryonic development, and the integrity of the organ. Deregulation of HSFs could be a risk factor for reproductive failure, cancer, neurogenesis, and neurodegenerative disorders (Ma, 2000). HSFY (heat shock transcription factor, Y chromosome) is a member of HSFs and has been located in the AZFb region of the Y chromosome (Rosenfeld, 2017). This gene is associated with spermatogenic failure, infertility (Peng, 2009; Zenteno-Ruiz, 2001), varicoceles (Meyfour, 2017), maturation arrest (MA) (McCann-Crosby, 2014), sertolicell-onlysyndrome (Meyfour, 2017) and plays a crucial role in the brain and cardiac development (Meyfour et al., 2017b).
SRY (sex determining region Y) encodes a transcription factor (a member of the high mobility group [HMG]-box family) (Jangravi et al., 2013) and involved in spermatogenesis, male sex determination, brain sexual differentiation, epigenetic processes, brain, cardiac and kidney development (Kamachi and Kondoh, 2013, Rosenfeld, 2017, Peng et al., 2009, Meyfour et al., 2017b).
Moreover, SRY and some SOX genes participate in carcinogenesis. Some diseases associated with SRY include 46, Xx / 46, XY Sex Reversal 1 (Zenteno-Ruiz et al., 2001), campomelic dysplasia (CD) (Foster et al., 1994), gonadal dysgenesis (Swyer syndrome) (Hunter et al., 2016); testicular regression syndrome (TRS)(Hunter et al., 2016), Turner's syndrome (Oliveira et al., 2009), infertility and azoospermia (Hamada et al., 1807), testicular (Kido and Lau, 2015) and prostate cancers (Ely et al., 2010), and brain disorders (Wu et al., 2009).
SRY expression and function in the human brain have been examined in several studies (Rosenfeld, 2017, Wu et al., 2009, Kido et al., 2017). SRY suppression in the human neuroblastoma cell line led to a down-regulation in enzymes involved in the dopamine synthesis of males and may describe the cause of more susceptibility of males in dopaminergic-based neurological disorders (Parkinson’s disease and schizophrenia) (Loke et al., 2015).
Although the SRY may be needed for the normal function of the male brain, an aberrant expression of this gene could impair neurogenesis and other disorders in mice pups (Kido et al., 2017). The SRY expression level in primed hESCs was higher than naive and embryoid bodies derived from hESCs (Taleahmad et al., 2019, Dehkordi et al., 2021 Apr). The results of the SRY knockdown in the primed cells showed upregulation of pluripotency markers and WNT pathway target genes in si-SRY treated cells and also increased the expression of several Y chromosome-linked genes in the primed cells (Taleahmad et al., 2019).
The RPS4Y1 encodes the S4 ribosomal protein, a part of the 40S subunit (Jangravi et al., 2013). This gene is associated with heart failure, turner syndrome, infertility, gonadoblastoma, prostate cancer, Parkinson’s disease, GVHD, and also participated in the brain and cardiac development (Meyfour et al., 2017b, Heidecker et al., 2010, Jangravi et al., 2013, Dasari et al., 2001, Fisher et al., 1990, Nielsen et al., 2010, Miklos et al., 2005, Sun et al., 2014).
Each of the target genes has various molecular and biological functions in different signaling pathways and human diseases (Table 1). Although, the precise mechanisms of functions have not been entirely understood. Most of the information that researchers identified are speculations based on the similarity of homologous regions to other genes. Gene Ontology (GO) of target genes is listed in Table 1.
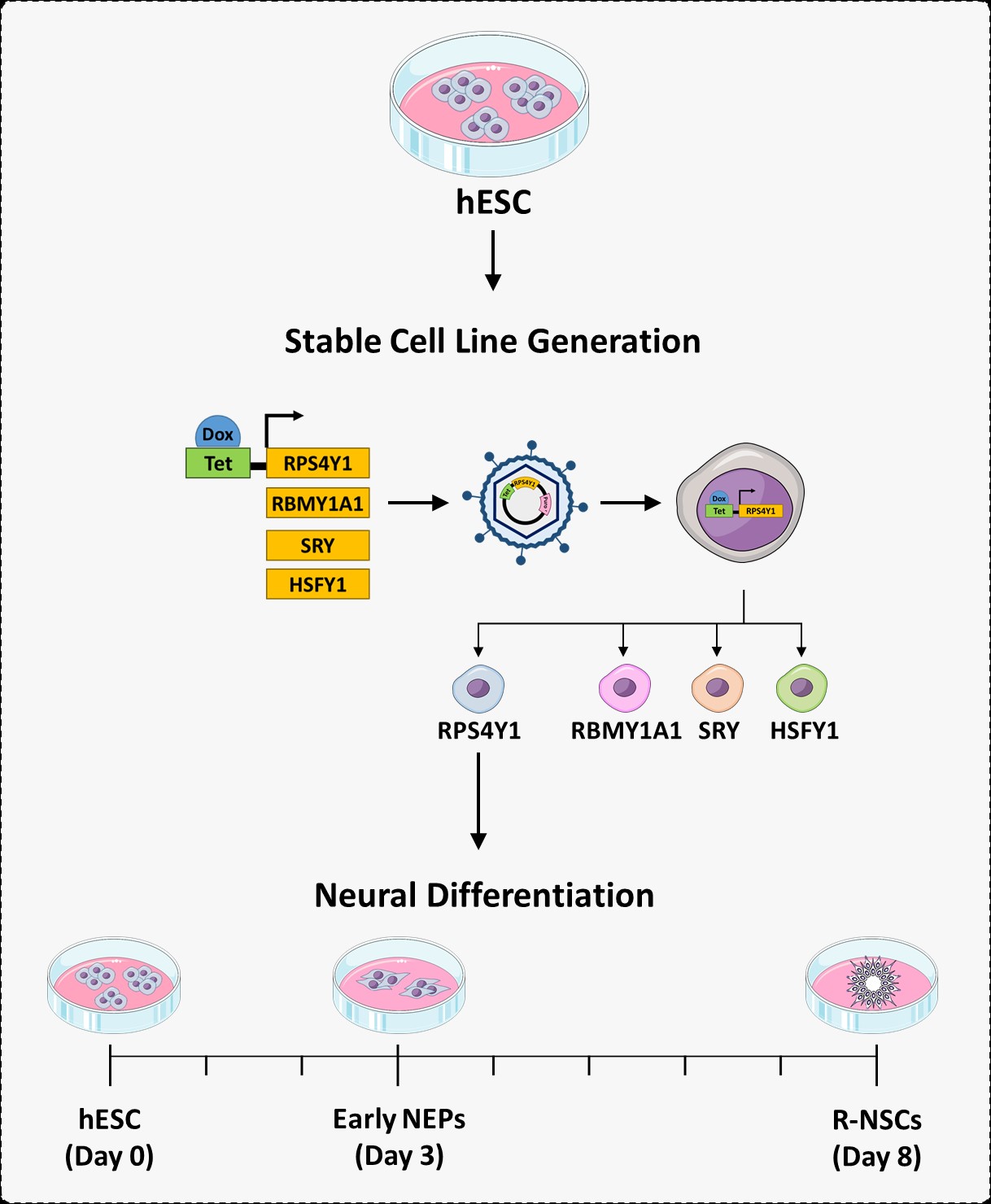
Based on the studies, we candidate HSFY1, SRY, RBMY1A1, and RPS4Y1 genes for more investigations (Skuse, 2000, Serajee and Mahbubul Huq, 2009, Lau and Zhang, 2000). To this end, we described development of the transgenic cell lines over-expressing target genes in an inducible manner. Then, they were characterized in terms of karyotype, pluripotency, and integrated gene expression level. We also discussed the applications of them in clinic and research.
In this study, we used a cell-based approach, using hESCs to generate four inducible cell lines able to increase gene of interest expression. As a proof of principle, we then differentiated one of these cell lines (RPS4Y1 over-expressing ESCs) to the neural stem cells.
RPS4Y1 is expressed during prenatal and infancy; indicating, it could play a critical role in early brain development (Meyfour et al., 2017b). Our results indicate a possible sex-dependent regulation of neural development that might underlie sexual dimorphism of human brain.
{kind=link}