OsATG8b restores autophagy activity in yeast scatg8 mutant
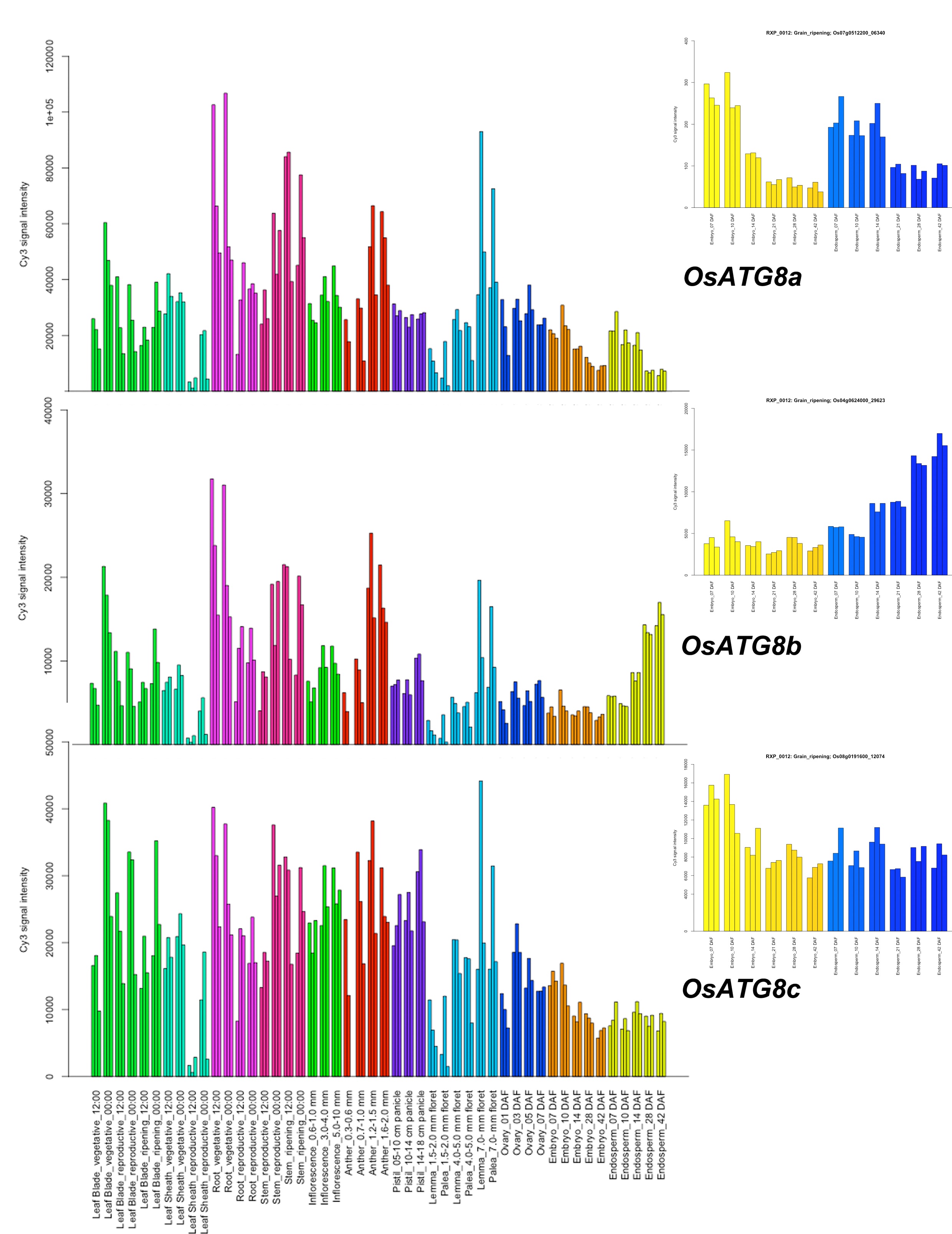
Six OsATG8s have been identified in the rice genome [16]. The ATG8 phylogenetic tree generated from amino acid sequences showed that plant ATG8s are clustered into two main subgroups. Subgroup I cover the most of the plant ATG8 family members, comprising OsATG8a, b and c. Subgroup II covers 1–3 plant ATG8 family members from each species, containing OsATG8d, e and f (Fig. S1). The existence of two subgroups may imply specific functions to each, besides possible redundancy.
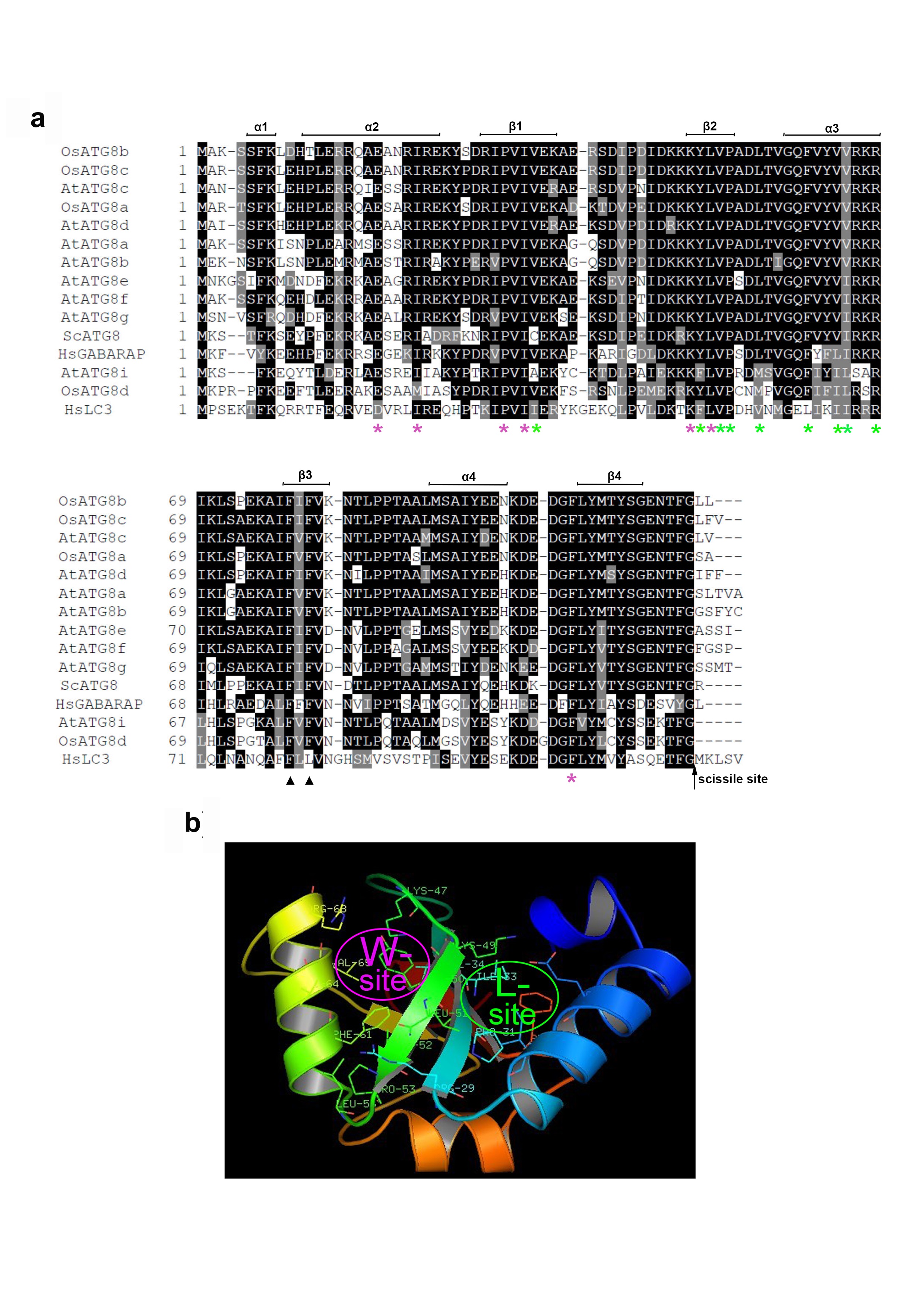
OsATG8b is encoded by a single gene (Os04g0642400) in rice. It is a soluble protein of 119 amino acids, with a predicted pI of 8.78. OsATG8b shares 81.8% amino acid identity with yeast ScATG8, 71.4% identity with human HsGABARAP, and 86.9% identity with Arabidopsis AtATG8a (Fig. S2a). Like other ATG8 proteins, OsATG8b has a conserved gly residue at the C-terminus for PE-conjugation (Fig. S2a). A 3D model prediction revealed that corresponds to the functional domains of yeast ScATG8, OsATG8b also contains a C-terminal ubiquitin-like domain and an N-terminal helical domain and two characteristic hydrophobic pockets named the W-site and the L-site (Fig. S2a and b) like yeast ScATG8 [36]. This implies that OsATG8b may have the autophagic function, similar to yeast ScATG8.
To verify the autophagic function of OsATG8b, we investigated whether OsATG8b rescues defects of ATG8-deficient (scatg8) yeast KVY5 [37]. OsATG8b cDNA containing the entire ORF was driven by the yeast GAL1 promoter in a plasmid (pYES260) and expressed in scatg8 yeast. OsATG8b can rescue the growth of the scatg8 yeast cells under N starvation (Fig. 1a). In yeast, the precursor amino-peptidase1 (prAPE1) was delivered to the vacuole for processing into mature APE1 (mAPE1) through the Cvt/autophagy pathway [19]. Thus, we monitored the protein levels of both prAPE1 and mAPE1 after 5 h of starvation. Both wild-type yeast and scatg8 cells complemented with OsATG8b accumulated mAPE1. In contrast, mAPE1 was detected in neither scatg8 cells nor the scatg8 cells transformed with the empty vector (Fig. 1b). This suggests that prAPE1 was delivered to the vacuole and processed to mAPE1. These results confirmed the autophagy role of OsATG8b and showed that OsATG8b is a functional homologue of yeast ScATG8.
OsATG8b expression is induced by N- and C-starvation
To determine the spatial and temporal expression pattern of OsATG8b, we employed qRT-PCR and the GUS reporter assay to examine OsATG8b expression. qRT-PCR analysis showed that OsATG8b transcripts accumulated in all studied organs, including roots, stems, leaves, leaf sheaths and panicles at different growth stages (Fig. 2a). Consistent with these results, in the OsATG8b promoter-GUS analysis, GUS activity was predominantly detected in all of the above-mentioned rice organs, and section analysis showed that OsATG8b is expressed in all tested cells (Fig. S3), thus suggesting that OsATG8b is expressed ubiquitously. Notably, OsATG8b transcript levels were higher in roots and leaves of plants at 45 days after germination (DAG) than in those of plants at other growth stages. At 60 DAG, OsATG8b transcript was relatively abundant in stems, leaf and panicle (Fig. 2a). The expression level of OsATG8b was also examined under N deficiency and darkness treatment for C starvation, respectively (Fig. 2b, c). OsATG8b transcript level increased in response to both N deficiency and darkness treatments. When rice seedlings were subjected to the N-free treatment, the expression level of OsATG8b gradually increased, peaking at 10 d after treatment application. Similarly, darkness treatment rapidly induced a rough three-fold increase in OsATG8b expression within 2 d after treatment. Taken together, these results suggest that OsATG8b may play a crucial role in regulating multiple developmental processes and in response to nutrient stresses.
GFP-OsATG8b is localized to autophagosomes
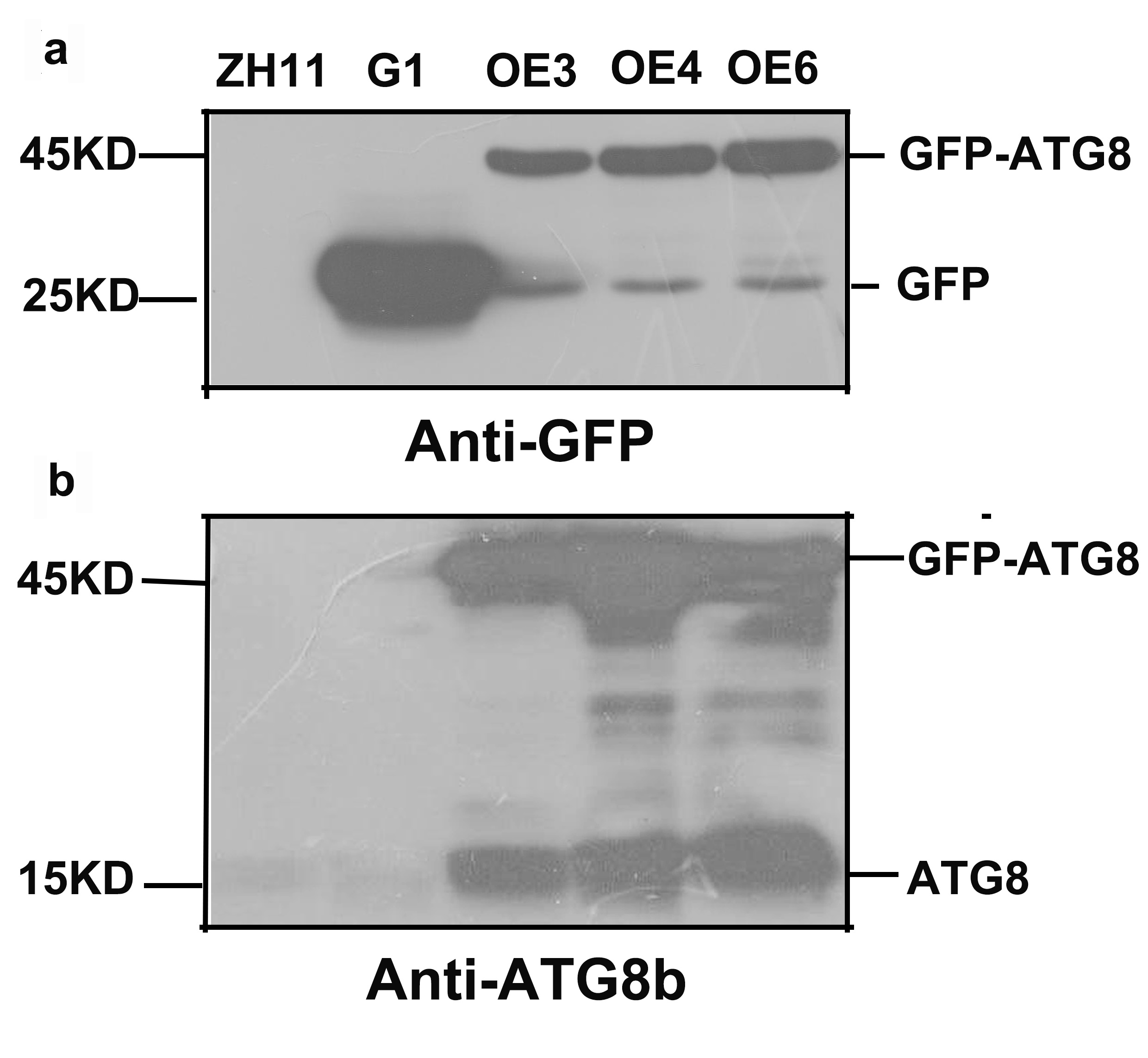
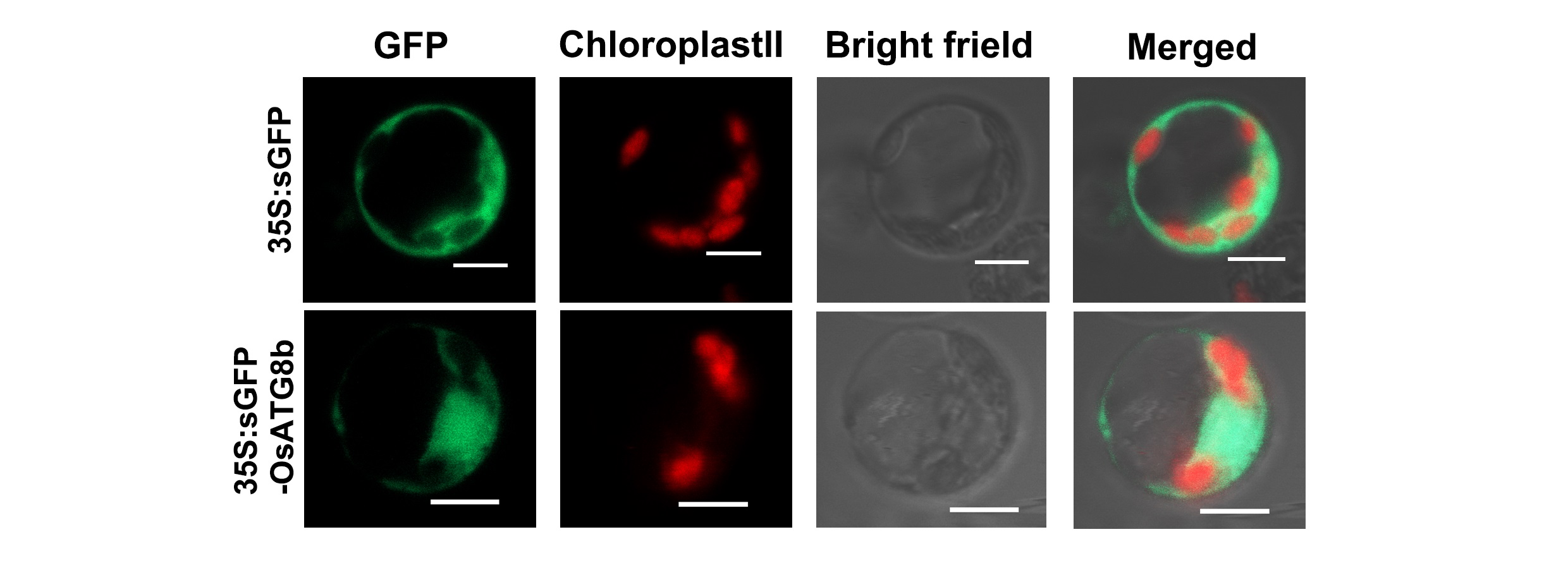
To determine whether OsATG8b is an autophagy marker for rice, GFP was fused to its N-terminus and transformed into scatg8 yeast cells. Under control conditions, GFP-OsATG8b was mainly localized to the cytosol with punctate distribution, whereas after starvation it accumulated within the vacuole of yeast (Fig. 3a). These data suggest that OsATG8b may be localized to the autophagosomes of cytosol in the control conditions and translocate from the cytosol to the vacuole in an autophagy-dependent manner after starvation in yeast. To further verify the above result in rice, sGFP-OsATG8b was also transiently expressed in rice protoplasts, but the data showed that the sGFP-OsATG8b fusion protein was localized to the membrane, cytoplasm, and nucleus (Fig. S4), similar to the free sGFP control. To further confirm sub-cellular localization, transgenic rice expressing sGFP-OsATG8b were generated under control of 35S promoter (Fig. 3b). The 5mm roots from tip were cut and immediately observed by LSCM. In sGFP-OsATG8b, GFP fluorescence was detected in the cytoplasm and nucleus; however, after 6h of incubation in the darkness with concanamycin A (an inhibitor of vacuolar H+-ATPase to help observation of autophagic bodies through increasing vacuolar pH [38, 39]), many vesicles with strong GFP signal and the spread of faint GFP signal were observed (Fig. 3b). These results indicate that the sGFP-OsATG8b-labeled puncta located in autophagosomes and the sGFP-OsATG8b can be used to visualize the progression of autophagy in rice, and overexpression of OsATG8b could increase the autophagic activity. Immunoblot analysis using proteins isolated from either ZH11 or transgenic sGFP or sGFP-OsATG8b rice plants showed that the OsATG8b antibodies recognized the endogenous as well as the GFP fusion proteins (Fig. S5). Meanwhile, we performed a sGFP-ATG8 processing assay by the levels of free GFP moiety in anti-GFP immunoblots. The results showed the ATG8 has already conjugated onto the autophagosome membrane and is able to be completed or delivered into the vacuole (Fig. S5).
OsATG8b affects root growth at grain germination
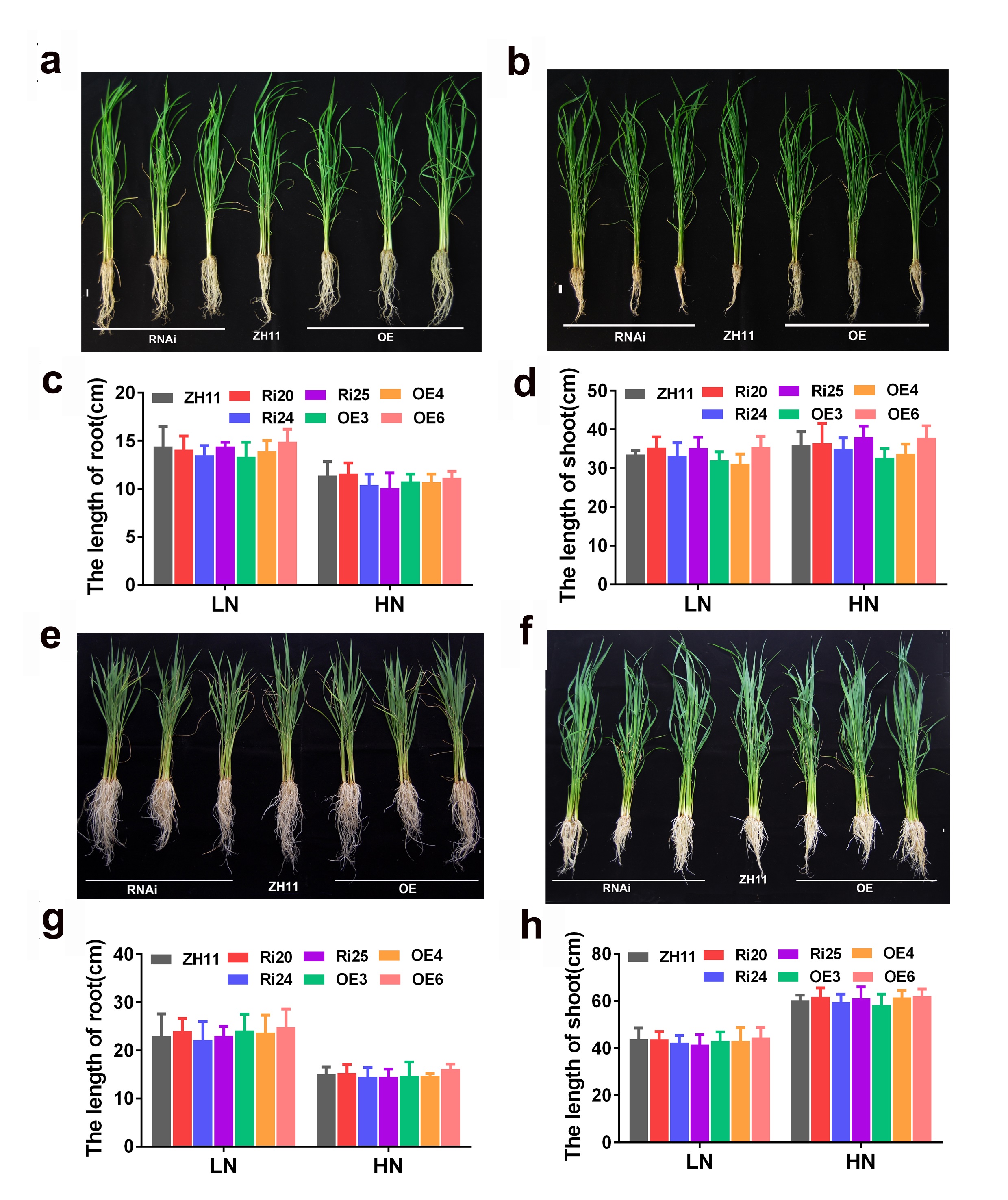
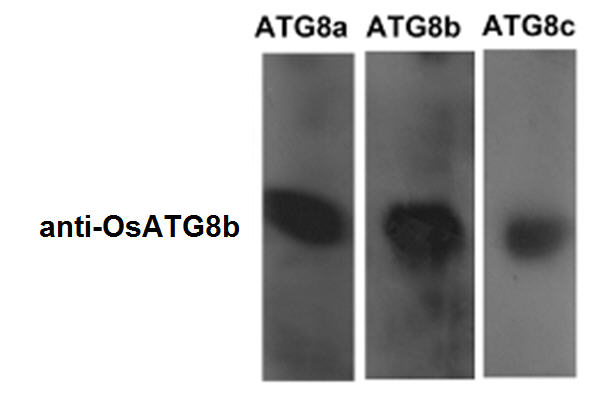
To further investigate the function of OsATG8b, OsATG8b over-expression (OsATG8b-OE) and RNA-interference (OsATG8b-RNAi) transgenic lines were generated. RT-PCR analysis showed that OsATG8b expression increased in OsATG8b-OE lines and reduced in OsATG8b-RNAi lines (Fig. 4a, b). The OsATG8b-RNAi construct was targeted specifically to the non-conserved 5’ end of OsATG8b outside the ubiquitin domain to avoid interference with other OsATG8 proteins. Three of the OsATG8b-RNAi lines (Ri20, Ri24 and Ri25) and three of the OsATG8-OE lines (OE3, OE4 and OE6) were selected for subsequent analysis. In order to observe the effect of altered OsATG8b expression to OsATG8a and OsATG8c, we detected the expression of OsATG8a and OsATG8c in the shoots and roots of the transgenic rice seedling at four leaf stage. The results showed there is no significant difference of OsATG8a or OsATG8c transcript level among ZH11, the OsATG8-OE lines or OsATG8b-RNAi lines (Fig. S6). To confirm whether autophagic activities is altered in the OsATG8b-RNAi and OsATG8b-OE lines, we examined the ATG8 autophagic activities in OsATG8b-RNAi, OsATG8b-OE and ZH11 lines using OsATG8b antibodies (Fig. 4c). The immunoblot analysis showed the expression levels of ATG8-PE (representing the forming or completed autophagosomes) and cytosolic ATG8 form were remarkably increased in OsATG8b-OE lines compared with ZH11 lines, and the quantified results showed there is slight decrease (about 17-20%) of them in OsATG8b-RNAi comparing with that in ZH11. These results indicated the autophagic activity is higher in OsATG8b-OE lines and a little lower in OsATG8b-RNAi than that in ZH11. In addition, we observed that the roots of OsATG8b-RNAi line were much shorter than those of ZH11 and OsATG8b-OE lines (Fig. 4d, e) at 7 DAG when germinated in water. To reveal how N affects autophagy in rice, growth of OsATG8b-RNAi and OsATG8b-OE lines were measured under low (0.2 mM N) and high (5 mM N) N levels. Under low and high N levels, the OsATG8b-RNAi and OsATG8b-OE lines appeared relatively normal phenotype and exhibited similar growth rate when compared with ZH11 at 30 or 60 DAG (Fig. S7). Neither root nor shoot length showed any significant difference among these lines (Fig. S7).
OsATG8b affects grain yield and grain quality in rice
The phenotypes of OsATG8b-RNAi and OsATG8b-OE rice at the reproductive stage were investigated in the paddy field under normal N conditions. Previous studies have shown that the autophagy-defective rice mutant osatg7 displayed complete sporophytic male sterility. However, OsATG8b-RNAi and OsATG8b-OE plants produced healthy pollen grains and could be fertilized normally. The statistical results showed that grain number and grain yield per plant increased in OsATG8b-OE plants but decreased in OsATG8b-RNAi ones, compared with ZH11 plants (Fig. 5). These data indicate that OsATG8b may be involved in grain development and yield.
The grains of OsATG8b-RNAi are brown-spotted hull and conatin chalky endosperm (Fig. 6a, b). This showed that it produced poor quality seeds. The percentage of hulled rice with chalkiness was higher in OsATG8b-RNAi lines compared to ZH11 (Fig. 6c). SEM revealed that there are many loosely packed and small starch granules in endosperm of OsATG8b-RNAi, which differed from the large and tightly packed starch granules in ZH11 (Fig. 6d). Conversely, endosperm starch granules of OsATG8b-OE and ZH11 grains seemed larger and tighter (Fig. 6d). Compared with ZH11, soluble protein content in OsATG8b-RNAi lines was lower while that in OsATG8b-OE lines was higher (Fig. 6e). However, starch content showed no significant difference among those lines (Fig. 6f).
OsATG8b affects N recycling to grains
To investigate whether OsATG8b plays a role in N recycling to grains in rice, we performed a pulse-chase assay with 15NO3-, as previously conducted with Arabidopsis [23, 40]. 15N and the 14N/15N ratio were measured (Fig. 7a). Plant DW was higher in OsATG8b-OE lines and lower in OsATG8b-RNAi lines than in ZH11 (Fig. 7b). This is similar to what was observed in Arabidopsis mutants (atg5, atg7) [13, 23]. HI, an important productivity indicator [41], was lower in OsATG8b-RNAi lines, but higher in OsATG8b-OE lines than in ZH11 (Fig. 7c), which shows that autophagy plays an important role at the grain filling stage.
NHI is a main index of the efficiency of N distribution to grains and N grain filling [23]. The NHI of OsATG8b-RNAi was lower than that of ZH11, while that in OsATG8b-OE as higher (Fig. 7d). As the NHI/HI ratio is considered a good indicator of NUE in plants [40], we then measured the NHI/HI ratio of OsATG8b-RNAi, OsATG8b-OE and ZH11. The results showed that the NHI/HI ratio increased dramatically in OsATG8b-OE lines, but decreased in OsATG8b-RNAi lines when compared to ZH11 (Fig. 7e). These data indicate that OsATG8b-mediated autophagy plays a role in grain NUE.
On the 7th d after 15NO3-labeling, 15N contents of OsATG8b-RNAi, OsATG8b-OE and ZH11 showed no significant differences. This is consistent with the normal growth of OsATG8b-RNAi and OsATG8b-OE lines under N-rich conditions (Fig. S8). The abundance of 15N in grains and remains were determined using isotopic ratio mass spectrometry, enabled us to calculate the partitioning of 15N in grains (15NHI) by combining these values with DW and N% data. 15NHI and the 15NHI:HI ratio, an indicator for NRE, were lower in OsATG8b-RNAi lines and higher in OsATG8b-OE lines than in ZH11 (Fig. 7f, g). Taken together, these 15N partitioning results show that OsATG8b-mediated autophagy significantly affects NRE during the grain filling stage.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}