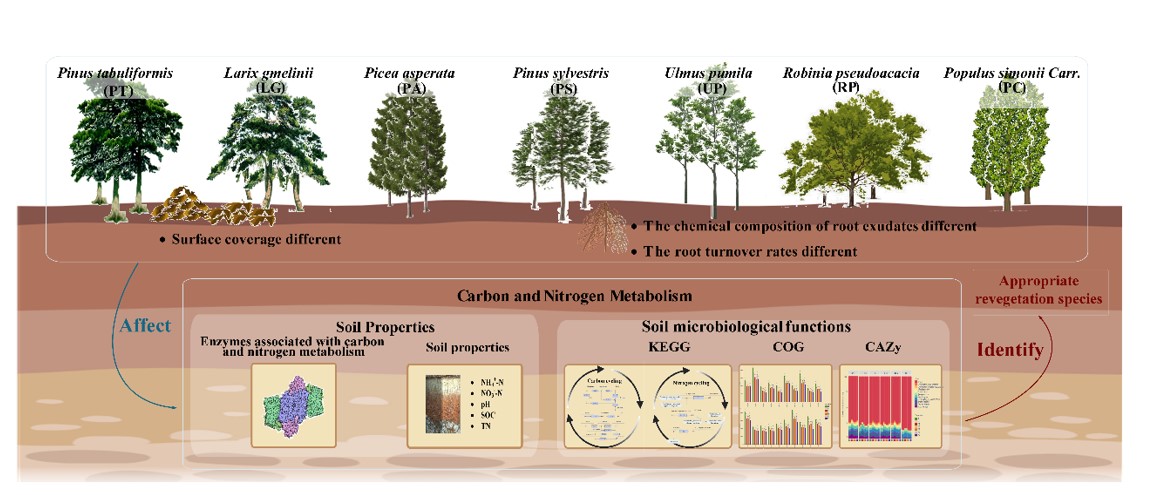
Soil properties and enzyme activities
The content of SOC (9.595 ± 0.091g/kg) was significantly higher in the soil of PT than in other soils (P < 0.05; Table 2). NO3−-N content differed significantly in 7 vegetation soils, which was highest in the soil of UP (60.425 ± 0.108 mg/kg; P < 0.05; Table 2). The soil of LG has the highest NH4+-N content (10.613 ± 0.061 mg/kg), but the NO3−-N content is the lowest (22.6 ± 0.059 mg/kg; P < 0.05; Table 2).
Table 2
Physical and chemical properties of soil under 7 types of vegetation
| |
NH4+-N
(mg / kg)
|
NO3−-N
(mg / kg)
|
pH
|
SOC
(g / kg)
|
TN
(%)
|
Bulk density(g/ml)
|
|
RP
|
9273 ± 0179abcd
|
56424 ± 0101c
|
7787 ± 0013bc
|
5227 ± 0303d
|
1094 ± 0059ab
|
04 ± 001b
|
|
LG
|
10613 ± 0061a
|
226 ± 0059g
|
779 ± 0008bc
|
6477 ± 0229c
|
0923 ± 0215b
|
042 ± 006ab
|
|
PA
|
8194 ± 0117d
|
4689 ± 0089d
|
786 ± 0011b
|
7719 ± 0136b
|
0489 ± 0112c
|
046 ± 002ab
|
|
PC
|
10146 ± 0187ab
|
32062 ± 0093f
|
7717 ± 0014c
|
7669 ± 0152b
|
1174 ± 0064a
|
043 ± 001ab
|
|
UP
|
8489 ± 0153cd
|
60425 ± 0108a
|
7997 ± 0003a
|
5307 ± 0198d
|
0930 ± 0036b
|
046 ± 004ab
|
|
PT
|
8883 ± 0146bcd
|
38575 ± 0125e
|
794 ± 0004a
|
9595 ± 0091a
|
0966 ± 0059b
|
05 ± 002a
|
|
PS
|
9789 ± 0212abc
|
58823 ± 0205b
|
786 ± 001b
|
6731 ± 0022c
|
0624 ± 0048c
|
044 ± 002ab
|
| Data are means ± SD in parentheses and different letters in a column indicate significant differences (Turkey's test, P < 005) SOC, total soil organic carbon; TN, total nitrogen; RP: Robinia pseudoacacia; LG: Larix gmelinii; PA: Picea asperata; PC: Populus simonii Carr; UP: Ulmus pumila; PT: Pinus tabuliformis; PS: Pinus sylvestris. |
The activity of N-acetylglucosaminidase was significantly different among 7 vegetation soils (P < 0.05; Table 3). The activities of β-glucosidase (170.776 ± 0.994 U/g) and alkaline protease (0.281 ± 0.005 U/g) were significantly highest in the soil of PC (P < 0.05; Table 3). Meanwhile, β-glucosidase and urease all showed significant minima (P < 0.05; Table 3) in the soil of RP, with a minimum of 56.799 ± 0.586, and 58.433 ± 1.868 U/g, respectively.
Table 3
Enzyme activity in soils under 7 types of vegetation
| |
C cycle-related enzymes (U/g)
|
N cycle-related enzymes (U/g)
|
| |
Dehydrogenase
|
β-glucosidase
|
Cellulase
|
Urease
|
Alkaline protease
|
N-acetylglucosaminidase
|
|
RP
|
0767 ± 0018b
|
56799 ± 0586e
|
17355 ± 0514e
|
58433 ± 1868e
|
0236 ± 0001b
|
8633 ± 0096e
|
|
LG
|
1475 ± 0072a
|
101816 ± 12c
|
23055 ± 0388b
|
246773 ± 0626c
|
0196 ± 0001c
|
955 ± 0022d
|
|
PA
|
1425 ± 0048a
|
116024 ± 0951b
|
30486 ± 0239a
|
300372 ± 193b
|
0189 ± 0002c
|
16699 ± 0035a
|
|
PC
|
0108 ± 0011c
|
170776 ± 0994a
|
34524 ± 1756ab
|
252935 ± 0278c
|
0281 ± 0005a
|
12377 ± 0041b
|
|
UP
|
1483 ± 0026a
|
93618 ± 1998cd
|
2216 ± 0457b
|
178035 ± 1167d
|
0198 ± 0002c
|
7972 ± 0029f
|
|
PT
|
1283 ± 0016a
|
90864 ± 0685d
|
20495 ± 0125b
|
170969 ± 0146d
|
0177 ± 0007c
|
11081 ± 0012c
|
|
PS
|
0208 ± 0008c
|
113722 ± 1307b
|
23825 ± 048b
|
378592 ± 0226a
|
0232 ± 0b
|
10719 ± 0027c
|
| Data are means ± SD in parentheses and different letters in a column indicate significant differences (Turkey's test, P < 005) RP: Robinia pseudoacacia; LG: Larix gmelinii; PA: Picea asperata; PC: Populus simonii Carr; UP: Ulmus pumila; PT: Pinus tabuliformis; PS: Pinus sylvestris. |
COG annotation based on metagenome sequencing
To determine the potential roles of microbial communities in the carbon and nitrogen cycle, the specific COGs involved in carbohydrate transport and metabolism, amino acid transport and metabolism were analyzed. The total items of carbohydrate transport and metabolism, and amino acid transport and metabolism were 1371 (6.70%) and 551 (9.23%) in the COG function classification, respectively. The significant differences of related COGs abundance in 7 vegetation soils were tested (one-way ANOVA, Turkey's test, P < 0.05), and the top 10 abundant COGs are selected and shown in Fig. 1 and Table S1.
Several carbohydrate-related functions enriched in the 7 samples of soil microorganisms were the major facilitator Superfamily COG0477 (33561 average reads), Dehydrogenase COG2133 (26861 average reads), and transporter activity COG1653 (26300 average reads). Furthermore, the abundance of COG0477, COG2133, and COG1653 was significantly higher in sample RP than that in other samples (P < 0.05). Regarding amino acid-related function, acetolactate synthase COG0028 (35191 average reads), succinyl-diaminopimelate desuccinylase activity COG0624 (35071 average reads), and peptidase s9 prolyl oligopeptidase active site domain protein COG1506 (26841 average reads) were dominated in the 7 samples and significantly highest in the sample RP (P < 0.05; Fig. 1b; Table S1).
A potential link exists between microbial taxa and their function of carbohydrate transport and metabolism, amino acid transport, and metabolism (Fig. S1). Fig. S1 showed that carbohydrates and amino acids in the 7 plant soils are predominantly transported and metabolised by the Solirubrobacter (Actinobacteria), Nocardioides (Actinobacteria), Bradyrhizobium (Proteobacteria), and Streptomyces (Actinobacteria).
Functional enzymes and their presentations in KEGG
To further investigate the differences in carbon and nitrogen cycling function under different vegetation types, we annotated genes to the KEGG enzyme. There were 21 enzymes (14 involved in carbon cycling and 7 involved in nitrogen cycling) involved in 17 KEGG pathways that have significant differences among 7 types of plant soils (P < 0.05; Fig. 2, 3). The detailed information for enzymes and KEGG pathways are provided in Tables S2. As one of the dominant enzymes involved in carbon cycling, β-glucosidase (EC3.2.1.21; average reads 13625) was involved in multiple carbon cycling pathways (Fig. 2,3; Table S2). Cellulose could be hydrolyzed into cellodextrin with endo-1,4-b-glucanase (EC3.2.1.4), followed by hydrolysis into cellobiose with endo-1,4-b-glucanase (EC3.2.1.4) and β-Glucosidase (EC3.2.1.21; Fig. 2a). Subsequently, cellobiose could be glycosylated into D-glucose with b-glucosidase (EC3.2.1.21). Meanwhile, β-D-Glucoside could be directly glycosylated into D-glucose with EC3.2.1.21(Fig. 2a). Except for EC3.2.1.21, α-Amylase (EC3.2.1.1; average reads 10798) and α-Glucosidase (EC3.2.1.20; average reads 10048) also dominated in enzymes involved in carbon cycling (Fig. 3a; Table S2). Dextrin and maltodextrin could be hydrolyzed into starch and maltose, respectively, by α-Amylase (EC3.2.1.1; Fig. 2a). Subsequently, maltose could be glycosylated into D-glucose with α-Glucosidase (Fig. 2a).
Regarding the 7 enzymes involved in nitrogen cycling, argininosuccinate lyase (EC4.3.2.1; average reads 7309), aspartate aminotransferase (EC2.6.1.1; average reads 4517), and acetylornithine deacetylase (EC3.5.1.16; average reads 5072) were dominated (Fig. 3b; Table S2). N-(L-Arginino) succinate could be hydrolyzed into fumarate with argininosuccinate lyase (EC4.3.2.1; Fig. 2b). N-Acetyl-L-citrulline could be hydrolyzed into citrulline with acetylornithine deacetylase (EC3.5.1.16; Fig. 2b). Compared with argininosuccinate lyase and acetylornithine deacetylase, aspartate aminotransferase was involved in several metabolisms. 2-oxoglutarate, 3-(4-Hydroxyphenyl) pyruvate and L-erythro-4-Hydroxyglutamate could be hydrolyzed into L-Glutamate, L-Tyrosine, and (4R)-4-Hydroxy-2-oxoglutarate, respectively, by aspartate aminotransferase (Fig. 2b).
A potential link exists between microbial taxa and enzymes involved in carbon and nitrogen cycling (Fig. S2). Fig. S1 showed that β-Glucosidase (EC3.2.1.21) and α-Glucosidase (EC3.2.1.20) in the 7 plant soils were produced by the Solirubrobacter (Actinobacteria), Nocardioides (Actinobacteria), Sphingomonas (Proteobacteria), and Streptomyces (Actinobacteria). α-Amylase (EC3.2.1.1) was produced by unclassified_c_Betaproteobacteria, Nocardioides (Actinobacteria), and Streptomyces (Actinobacteria). Different from enzymes involved in carbon cycling, the potential link between microbial taxa and several enzymes involved in nitrogen cycling has differences. Argininosuccinate lyase (EC4.3.2.1) was produced by unclassified_p_Acidobacteria, Nocardioides (Actinobacteria), and Streptomyces (Actinobacteria). Argininosuccinate lyase (EC4.3.2.1) was produced by unclassified_p_Acidobacteria, Nocardioides (Actinobacteria), and Streptomyces (Actinobacteria). Aspartate aminotransferase (EC2.6.1.1) was produced by Bradyrhizobium (Proteobacteria) and unclassified_p_Acidobacteria. Acetylornithine deacetylase (EC3.5.1.16) was produced by Bradyrhizobium (Proteobacteria), unclassified_p_Acidobacteria, Solirubrobacter (Actinobacteria), Streptomyces (Actinobacteria), and unclassified_p_Chloroflexi.
CAZyme annotation based on metagenome sequencing
To better understand the carbohydrate metabolism in the soil ecosystem with 7 vegetation types, we annotated our metagenome for genes encoding carbohydrate-active enzymes (Fig. 4). The relative abundance of glycoside hydrolases (GHs), glycosyltransferases (GTs), polysaccharide lyases (PLs), carbohydrate esterases (CEs), carbohydrate-binding modules (CBMs), and auxiliary activities (AAs) showed significant difference among 7 vegetation soils (P < 0.05; Fig. 4). GTs and GHs which involve in soil organic C decomposition and biosynthesis respectively had higher relative abundance (Relative abundance > 30%; Fig. 4e, f).
The relationship between microbial community composition and CAZyme-encoding genes was shown in Fig. 5. The abundance of CAZyme-encoding genes varied across the bacterial phyla. As shown in Fig. 5, most of these genes in the soil microorganisms under 7 vegetation types were from Nocardioides (Actinobacteria), Solirubrobacter (Actinobacteria), Bradyrhizobium (Proteobacteria), Streptomyces (Actinobacteria), Sphingomonas (Proteobacteria), Conexibacter (Actinobacteria), and Mycobacterium (Actinobacteria). Members of Actinobacteria, Acidobacteria, and Proteobacteria were dominant in carbohydrate-active modules of GHs, GTs, CEs, and CBMs, whereas those from Acidobacteria were dominant in PLs (Fig. 5).
Relationships between the abundance of microbial genes, soil properties and enzyme activities
The relationship between the soil properties and enzyme activities and the abundance of six microbial gene groups for C cycling showed that TN was significantly correlated with GHs, GTs, PLs, CEs, CBMs, and AAs (P < 0.05; Fig. 6b). In addition, NO3−-N showed a significant correlation with GHs which were involved in soil organic C decomposition (P < 0.05; Fig. 6b). Urease and dehydrogenase were significantly correlated with TN (P < 0.05; Fig. 6a). Meanwhile, N-acetylglucosaminidase showed a significant relationship with SOC and NO3−-N (P < 0.05; Fig. 6a). The relationship between the soil properties and enzyme genes involved in carbon-nitrogen cycling was shown in Fig. 6c. Interestingly, NO3−-N and pH showed a significant correlation (P < 0.05) with several enzymes involved in carbon cycling but have no significant relationship with enzymes involved in nitrogen cycling (P > 0.05; Fig. 6c).
{kind=link}