Drugs are an emerging class of environmental pollutants [19]. In recent years, the presence of drugs and their metabolites and transformation products in aquatic environments has become a matter of close concern because of the potential adverse effects of these compounds on organisms [20]. Various drugs have been reported to be detected in many environmental samples around the world, mainly in wastewater from sewage treatment plants, surface water, seawater, groundwater, and fish at concentrations ranging from ng/L to mg/L [21]. The presence of a large number of pharmaceutical contaminants in the aquatic environment poses a potential threat to the outcome of the aquatic biota and cannot be predicted at present [22]. Therefore, we urgently need to evaluate the potential effects of drugs on aquatic plants and animals to provide new insights into the aquatic environment.
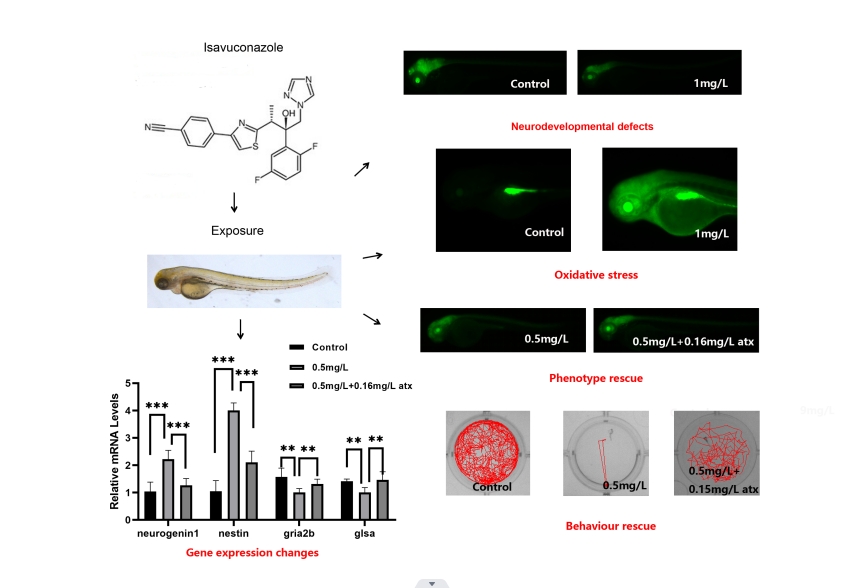
Zebrafish can help infer the potential effects and risks of drugs in their aquatic organisms and aquatic ecosystems [23]. In the present study, we evaluated the effects of the antifungal drug, isavuconazole, on neurotoxicity in larval zebrafish after exposure. Our results showed that isavuconazole exposure resulted in shortened body length, slowed heart rate, and reduced tail flick frequency in zebrafish larvae. At the same time, isavuconazole caused zebrafish oxidative stress response and nerve-related gene expression disorders, while astaxanthin could reduce the neurodevelopmental defects caused by isavuconazole-induced oxidative stress, reduce the level of ROS, and restore motor behaviour.
Studies have found that under normal physiological conditions, fish and other vertebrates can rapidly clear ROS through antioxidant enzymes, such as superoxide dismutase and catalase [24]. However, some high-dose clinical drugs can cause excessive generation of reactive oxygen species (ROS) in tissues, thereby destroying major cellular macromolecules and inducing oxidative stress leading to cell damage and apoptosis [25–26]. In this study, we detected the enzyme activity indicators of zebrafish larvae, including SOD enzyme (the first line of defence against oxidative damage), CAT enzyme (a key enzyme in the biological defence system) and MDA content (assessing peroxide). It was found that the enzyme activities of SOD and CAT increased, and the content of MDA decreased. At the same time, we also detected the activity of the AchE and ATPase enzymes related to the nervous system, and the results of the exposure group showed a decline in varying degrees [27]. We believe that its mechanism of action may be through the inhibition of ATPase on the synaptosomal membrane of aquatic organisms, resulting in the accumulation of neurotransmitters such as AchE on the postsynaptic membrane, and finally inhibiting the activity of brain AchE. It shows that isavuconazole exposure can affect the normal physiological function of zebrafish and even lead to the death of fish.
To further evaluate the toxicity of the drug, we analysed the mRNA expression levels of genes related to neurodevelopment and neurotransmitter transmission in zebrafish larvae treated with 72 hpf with reference to previous studies [28–29]. The expression levels of neurodevelopment-related genes such as GFAP, which are related to neurodevelopment, changed, and genes related to the neurotransmitter system also showed a downward trend in general. The results showed that they had been specifically damaged, affecting the development of the nervous system and resulting in a decline in its function.
Movement behaviour is also one of the important indicators to evaluate the neurotoxicity of zebrafish embryos [30]. After isavuconazole exposure, several important parameters of juvenile fish, such as total distance moved, time moved and speed of movement decreased. These data suggest that isavuconazole exposure can affect locomotor behaviour in zebrafish.
To further explore the underlying molecular mechanism of isavuconazole-induced neurodevelopmental deficits in zebrafish, we used astaxanthin (an oxidative stress inhibitor) to study the phenotypic changes after isavuconazole exposure. Previous studies have shown that astaxanthin has a protective effect on pamiparib-induced zebrafish neuronal cell damage, and astaxanthin can also rescue the neurodevelopmental deficits in zebrafish after propranolol exposure [31–32]. We found that likewise, astaxanthin was effective in reducing isavuconazole-induced oxidative stress, rescuing the enzymatic activity of total ATPase and AchE. In addition, we analysed whether ATX can rescue neurodevelopmental defects after exposure to isavuconazole. As expected, the expression of nerve-related gene mRNA also reached a level similar to that of the control group. Finally, we rescued isavuconazole-induced motor dysfunction in zebrafish with astaxanthin. The results showed that the length of behavioural trajectories was significantly increased after co-exposure to isavuconazole and astaxanthin compared to the isavuconazole-exposed group. Co-exposure to isavuconazole and astaxanthin also significantly increased exercise measures such as total exercise distance, exercise time, and average speed. These results further demonstrate that isavuconazole induces neurodevelopmental deficits and behavioural abnormalities in zebrafish mainly by modulating ROS-mediated oxidative stress signalling. This study provides new insights into the neurotoxicity and potential mechanism of isavuconazole in juvenile zebrafish. In the future, we should focus on deeper research on the mechanism and evaluate the toxic effect of isavuconazole from different perspectives.
{kind=link}