In the occurrence of skin disease, researchers observe microbial dysbiosis both in the gut and skin [10, 28]. For example, the microbiological composition of healthy skin is balanced with proper quantities of human and microbial antimicrobial peptides (AMPs). AMPs are a secretory product of epithelial and immune cells regulated by the Toll-like receptor (TLR) pathway. AMPs can maintain intestinal homeostasis and prevent the entry of gut bacteria into the bloodstream. When TLR pathways are inhibited, gut allostasis occurs. Dysbiosis of the gut microbiome influences the conversion of complex polysaccharides into vitamins and short-chain fatty acids (SCFAs) to improve the integrity of the gut barrier [10]. They may also affect nitric oxide (NO) production and influence blood flow through the denitrification pathway [29]. Impairment of intestinal epithelial cells leads to reduce production of AMPs and immunoglobulin A (IgA), which exacerbates gut microbial dysbiosis. As a result, gut bacteria enter the bloodstream through the gut wall and then reach the skin, causing dysbiosis of the skin microbiome and tissue inflammation [10, 11, 30].
The skin performs its functions and undergoes constant renewal during homeostasis. Gut microbial dysbiosis can cause skin allostasis [11, 31]. For example, the dysbiosis of Firmicutes and Bacteroides alters the serological cytokine levels and promotes inflammation, leading to the development of acne vulgaris [31]. This interaction is mainly mediated by the immune system [10, 32]. Additionally, bacterial metabolites, such as butyrate, are related to the integrity of the epithelial barrier which engages to protect the skin [10, 30, 33, 34]. Intact skin is crucial for maintaining homeostasis [35]. This intricate interaction is known as the gut–skin axis. However, there are few reports on the role of gut–skin axis in skin fibrosis [28]. Thus, we designed an MR analysis to determine the impact of the gut microbiome on the skin.
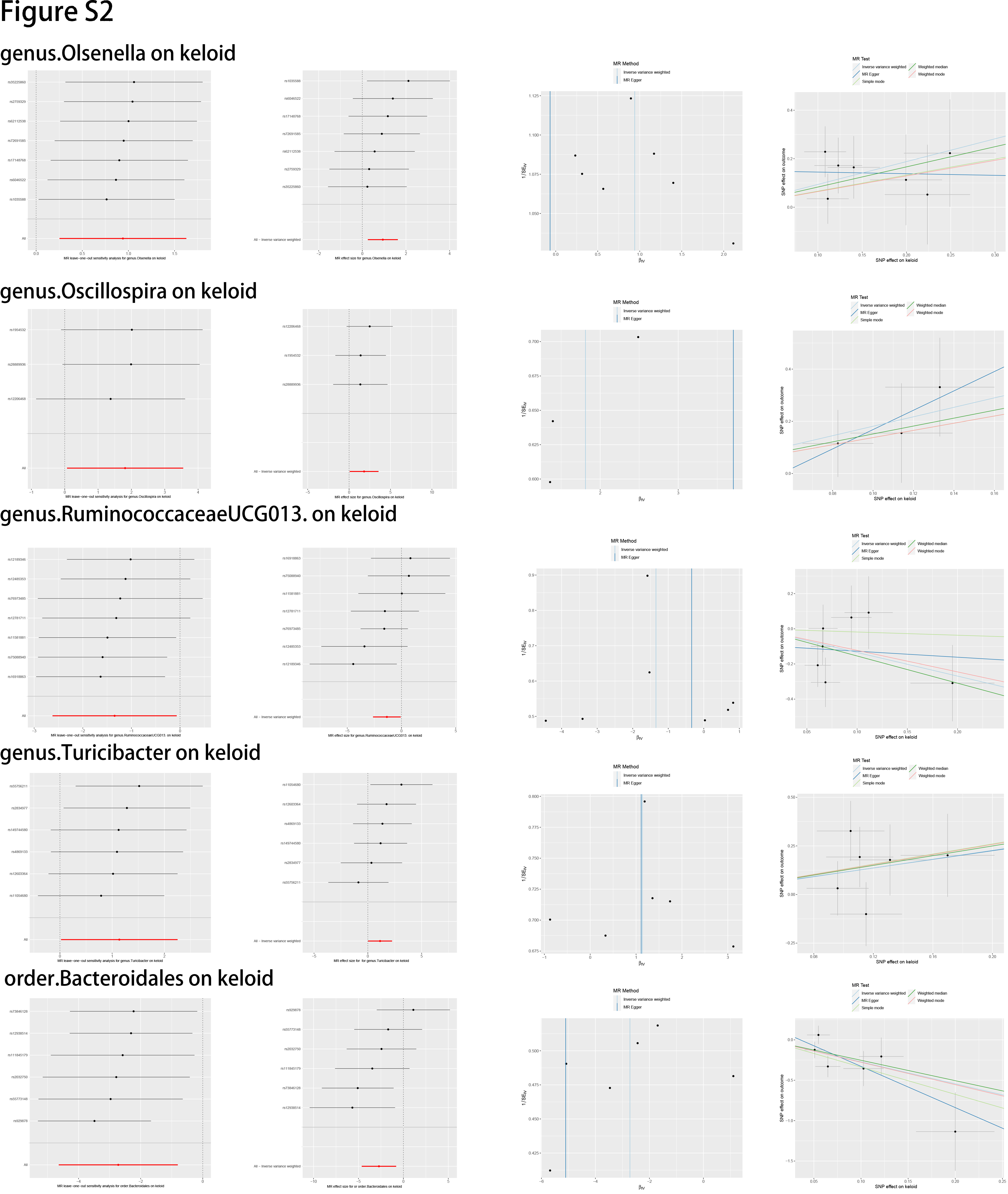
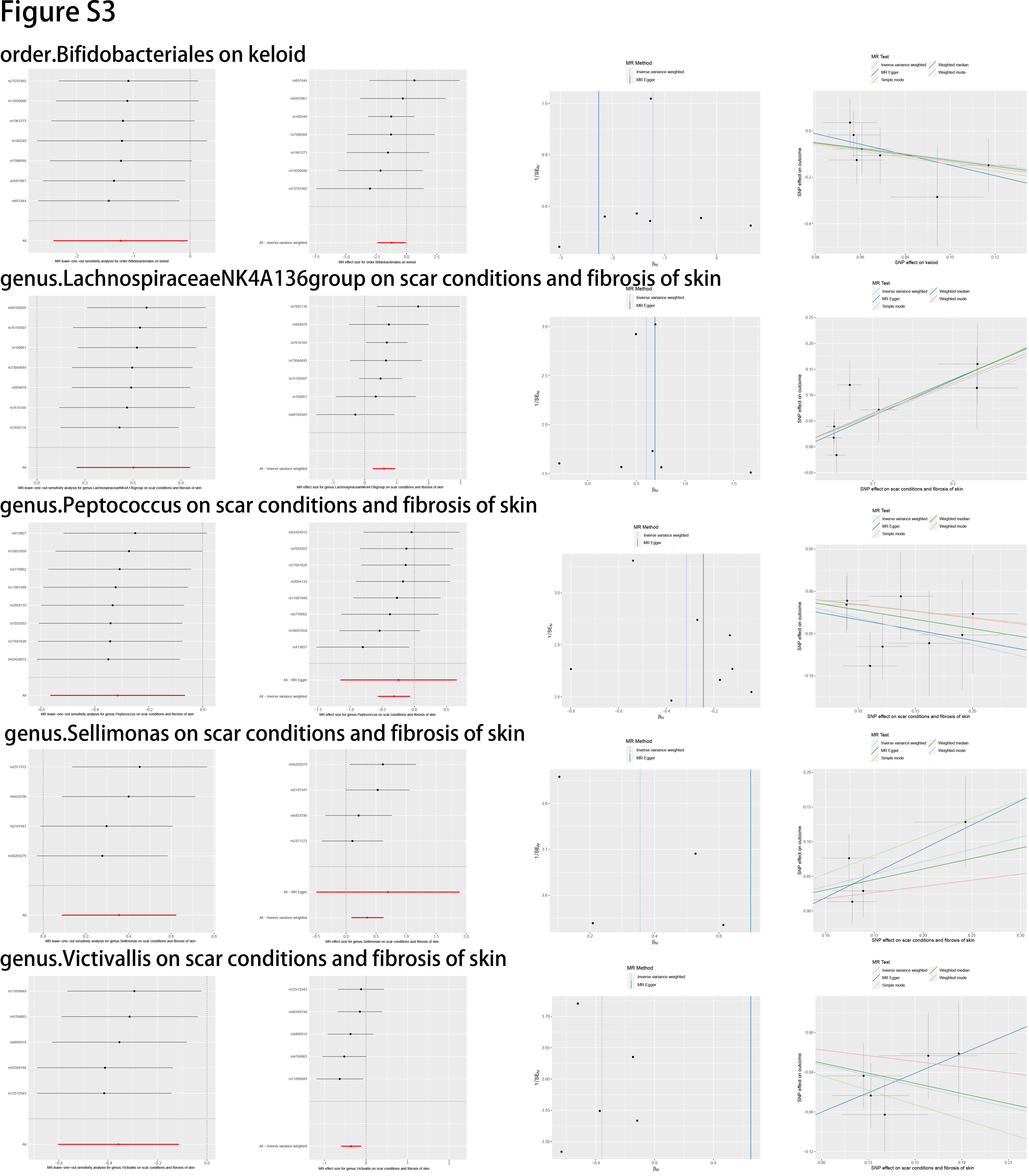
In this study, we evaluated the casual association between the abundance of specific bacterial signatures and the risk of skin fibrosis. Ten bacterial traits showed protective effects against skin fibrosis: Class Actinobacteria, Class Bacteroidia, family Bifidobacteriaceae, family Rikenellaceae, genus Lachnospiraceae (UCG004 group), genus Ruminococcaceae (UCG013 group), order Bacteroidales, order Bifidobacteriales, genus Peptococcus, and genus Victivallis. The genus Olsenella, genus Oscillospira, genus Turicibacter, genus Lachnospiraceae (NK4A136group), and genus Sellimonas are risk factors for skin fibrosis. In a randomised clinical pilot trial, participants consecutively consumed milk containing family Bifidobacteriaceae twice a day for 8 weeks. Compared with the pre-intake period, researchers found that the relative abundance levels of family Bifidobacteriaceae were significantly increased, and the skin condition of participants improved [36]. Genus Lachnospiraceae (UCG004 group) acts as probiotics and increases the production of SCFAs including butyric acid for skin homeostasis [33, 37]. Sodium butyrate dampens the profibrotic response induced by TGF-β1 in human dermal fibroblasts [38]. A study reported higher abundance levels of genus Turicibacter in patients with systemic lupus erythematosus than in healthy individuals [39]. These results are consistent with those of our study and support the existence of a gut–skin axis. Besides, Order Bacteroidales and genus Olsenella are associated with checkpoint blockade immunotherapy in melanoma [40, 41]. The mechanism underlying other bacterial traits in the gut–skin axis remains to be elucidated.
There are currently no effective treatments for skin fibrosis [2]. With the development of research on gut microbiome and skin fibrosis, we can elucidate the mechanisms of skin fibrosis and explore new therapeutic targets. These include the intake of probiotics, transplantation of the faecal microbiome, dietary modification, and drug–microbiome combination treatment [42–45]. Relative abundances of gut microbiome were regularly detected in the suspected population. Before the disease onset, early screening and diagnosis can be performed by engineering the gut microbiome and restoring intestinal homeostasis. In the context of skin fibrosis, we propose novel and effective therapeutic strategies based on changes in specific gut microbiome abundance levels. Based on these levels, we can evaluate the effects of treatment and adjust therapeutic strategies for precise treatment. After treatment, diet can be leveraged to optimise SCFA production, maintain a healthy gut–skin axis, and reduce the risk of recurrence [46]. To achieve this goal, we not only need appropriate data analysis to probe the causal association between the gut microbiome and skin fibrosis but also require a large number of rigorous clinical trials for validation.
Our study has some limitations. First, the results might not be entirely applicable to individuals of non-European descent because almost all samples are European, and only a few gut microbiome samples are from other races [17, 20, 21]. Second, there might be a possibility of identifying false-positive findings owing to the absence of no Bonferroni correction. However, the results with IVW-derived P values less than 0.05 should also be treated cautiously [14]. Third, the GWAS data for the gut microbiome were coordinated using by 16S rRNA gene sequencing, and the lowest taxonomic level was the genus. As a result, it is difficult to estimate the relationships between specific strains or species and skin fibrosis [17, 18]. Additionally, we cannot exclude a reverse causal association between the gut microbiome and skin fibrosis because of the relatively small size of the gut microbiome. We hope that future GWAS data of the gut microbiome will be more sufficient, and we can perform analyses between a specific species and skin fibrosis in both European and non-European populations to reduce bias and improve universality.
{kind=link}
{kind=link}
{kind=link}