Endozoic and tank water bacterial communities
Altogether, 50 endozoic and water samples (plus one negative control) were sequenced, and 7,110,067 high quality sequences were obtained (median frequency per sample was 107,522, min. 2, max. 911,443). Denoising yielded a total of 11,105 ASVs. After filtering mitochondrial and chloroplast sequences, the number of ASVs decreased to 10,946. Forty-three samples yielded enough high-quality reads (at minimum sequencing depth above 10,000) for downstream analyses (19/21 cloacal, 16/20 oral, and 8/9 tank water samples).
The highest number of observed features (OF) and phylogenetic diversity (Faith’s PD) was found in tank water samples (median OF = 603, IQR = 450; median Faith’s PD = 42.8, IQR = 53.9), followed by oral samples (median OF 473, IQR = 428; median Faith’s PD 40.2, IQR = 26.2), and cloacal samples (median OF = 308, IQR = 235; median Faith’s PD = 28.4, IQR = 20) (Fig. 2a). ASV richness and evenness (Shannon’s index) was highest in oral samples (median = 6.28, IQR = 1.33), followed by cloacal (median = 5.62, IQR = 1.66), and tank water samples (median = 5.44, IQR = 1.67) (Fig. 2b). Alpha diversity showed significant differences among sample sites (Kruskal-Wallis rank sum test) for OF and Faith’s PD (p-value < 0.01), while pairwise sample site comparisons (Wilcoxon rank sum test) showed differences between cloaca vs. oral samples and cloaca vs. tank water (q-value < 0.05). Shannon’s index showed weak statistical difference among samples sites (p-value = 0.04). Pearson correlation on alpha diversity indices against size of the turtle (CCL) in individual sample site groups showed strong negative correlation between cloacal samples and OF and Faith’s PD (R = -0.67, p-value = 0.002 and R = -0.59, p-value = 0.007, respectively) (Fig. 2c), while Shannon’s index showed weaker negative correlation with CCL (R = -0.53, p-value = 0.021). Oral and tank water alpha diversity did not show any correlation effects. Further, within cloacal samples, differences were detected (Kruskal-Wallis p-value < 0.05) in age range (OF, pairwise Wilcox: adult vs. juvenile q-value = 0.024; Faith’s PD, pairwise: adult vs. juvenile q-value = 0.024) and sex of the turtles (OF, pairwise Wilcox: male vs. ND q-value = 0.004; Faith’s PD, pairwise: male vs. ND q-value = 0.018), which are directly related to turtles’ size.
Bacterial communities among samples sites differed based on PERMANOVA for Bray-Curtis (R2 = 0.09, Pr(> F) = 0.001), Jaccard (R2 = 0.08, Pr(> F) = 0.001), unweighted and weighted UniFrac (R2 = 0.12, Pr(> F) = 0.001; R2 = 0.13, Pr(> F) = 0.001, respectively), and robust Aitchison distances (R2 = 0.24, Pr(> F) = 0.002). Pairwise PERMANOVA detected differences between all sample site pairs for most beta diversity metrics except robust Aitchison where only cloaca vs. tank water and tank water vs. oral samples pairs were significantly different (Pr(> F) ≤ 0.002). Highly ranked ASVs impacting the distribution of samples on rPCA biplot were assigned to Gammaproteobacteria, order Oceanospirillales, Shewanella algae, Vibrio sp., and NS3a marine group (Fig. 2d). Within cloacal samples, significant differences in bacterial communities were detected between adults vs. juveniles (Bray-Curtis, Jaccard, and unweighted UniFrac Pr(> F) < 0.05) and ND vs. males (all distances except robust Aitchison Pr(> F) < 0.05) or females (Jaccard and weighted UniFrac Pr(> F) < 0.05). In oral samples there were differences between early vs. late hospitalization duration (all distances except robust Aitchison Pr(> F) < 0.05) and early vs. mid (weighted UniFrac Pr(> F) < 0.05) hospitalization durations.
Cloacal bacterial communities differed between age groups based on PERMANOVA for Bray-Curtis (R2 = 0.15, Pr(> F) = 0.018), Jaccard (R2 = 0.14, Pr(> F) = 0.008) and unweighted UniFrac distance (R2 = 0.16, Pr(> F) = 0.013). Pairwise PERMANOVA showed significant differences only between adults and juveniles for all above-mentioned diversity measures (Pr(> F) ≤ 0.009). The differences between adults and juveniles are observed in community composition and structure as well: phylum Verrucomicrobiota appears more often in juveniles, juveniles also have higher RA of Rhodobacterales (Alphaproteobacteria), Cardiobacterales (Gammaproteobacteria), Kineosporiales (Actinobacteriota), Oligoflexales (Bdellovibrionota), Arcobacteraceae (Campylobacterales), and more Flavobacteriales (Bacteroidota) than adults. On the other hand, adults carry more Bacteroidales (Bacteroidota), Pasteurellales (Gammaproteobacteria), Helicobacteraceae and Campylobacteraceae (Campylobacterales), and Leptotrichiaceae (Fusobacteriales). The low number of samples (≤ 10) in each age group prevented us from further differential abundance analyses. Detailed taxonomic composition of bacterial communities found in oral, cloacal and tank water samples is reported in Supplementary Results, Table S3 with additional visualizations available at Github.
Differential abundance analysis detected 33 significantly DA features (post adjusting for multiple testing) out of which two were differentially abundant in cloaca (ASV467 Rhodobacteraceae and ASV4007 Shewanella algae), and three in oral samples (ASV9794 Truepera sp., ASV6166 and ASV467 both belonging to Rhodobacteraceae) (Fig. 3a). Tank water had 28 DA ASVs when tested against cloaca or oral samples, most of which belonged to typical marine taxa (Cryomorphaceae, NS3a marine group, SAR406 clade, Rhodobacteraceae, Nitrincolaceae, etc.). When collapsed to species level, 75 significantly DA taxa were detected, out of which 13 in cloaca and 15 in oral samples (Fig. 3b). Depending on the tested sample site pair several features would be DA in both oral and cloacal samples when compared to tank water e.g., genera Marinifilum, Tenacibaculum, and Labrenzia, members of Cardiobacteriaceae and Comamonadaceae families (Fig. 3b).
Endozoic and tank water fungal communities
Overall, 29 samples (plus one negative control) were sequenced, and 2,208,729 high quality sequences were obtained (median frequency per sample was 69,203, min. 11,722, max. 150,142). Denoising yielded a total of 9,547 ASVs. Based on alpha rarefaction curves, 27 samples yielded enough high-quality reads (at minimum sequencing depth at 25,000 reads) for downstream statistical analyses requiring rarefied data (19/20 cloacal, and 8/9 tank water samples). All samples except negative control were used in compositional data analyses.
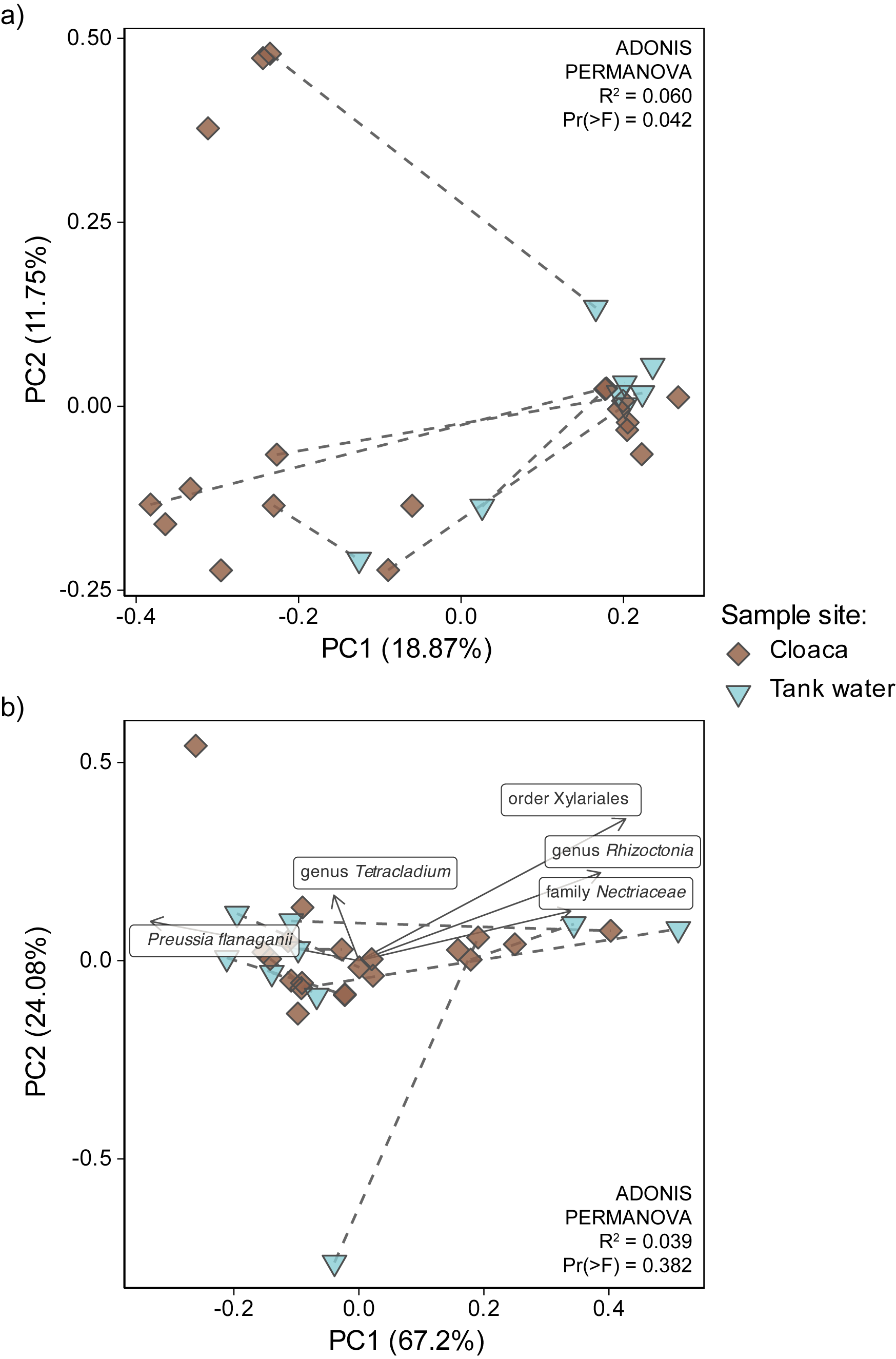
Tank water samples had significantly higher number of observed features and phylogenetic diversity (median = 674, IQR = 97.8; Faith’s PD median = 98.0, IQR = 10.5) than cloacal samples (median = 509, IQR = 372, Faith’s PD median = 66.6, IQR = 51.7) (Fig. 4a) based on Kruskal-Wallis rank sum test (OF p-value = 0.008; Faith’s PD p-value = 0.007). Shannon’s diversity was not detected as significantly different between sample sites (tank water median = 8.66, IQR = 1.31; cloaca median = 6.76, IQR = 2.58) (Fig. 4b). There were no significant differences in alpha diversity values within cloacal or tank water fungal communities when tested for age range, sex, or rehabilitation duration. The only beta diversity metric that showed significant differences between tank water and cloaca was unweighted UniFrac (R2 = 0.06, Pr(> F) = 0.042). Unweighted UniFrac PCoA showed unclear sample groupings, however, there is a separation of cloacal and corresponding tank water samples on PCA1 axis (Fig. S1a, corresponding samples connected with dashed lines). Compositional data analysis rPCA did not show clear separation of samples sites, and top ranked ASVs belonged to Preussia flanaganii, genus Tetracladium, order Xylariales, genus Rhizoctonia, and family Nectriaceae (Fig. S1b). When collapsed to species level, cloacal and tank water samples each had five differentially abundant fungal taxa detected. In cloacal samples DA taxa were Leptodiscella, Elaphomyces, Sclerocleista, Gliomastix, Thelephora; and in tank water they were Cladosporium, unidentified Leotiomycetes, unidentified Sordariomycetes (Chaetosphaerilaes), Nectria, and Humicola - although none of them were statistically significant after correction for multiple testing (Fig. 4c). Cloacal and tank water fungal communities were represented mostly by phyla Ascomycota (average RA ± standard deviation in cloaca and tank water = 57 ± 15% and 53 ± 9%, respectively), Basidiomycota (19 ± 9% and 22 ± 3%), Glomeromycota (7 ± 6% and 10 ± 12%), Mortierellomycota (2 ± 2% and 3 ± 2%) unidentified Fungi (13 ± 20% and 11 ± 7%) (Fig. 4d). Detailed taxonomic composition of fungal communities found in cloacal and tank water samples is reported in Supplementary Results and Table S4.
Epizoic and endozoic bacterial communities
After trimming epizoic, endozoic, and tank water sequences to V4 region of 16S rRNA gene (65 samples in total) and denoising, 10,072,866 high quality sequences were merged across all sequencing events (median frequency per sample was 126,629, min. 0, max. 946,183) and yielded 13,263 ASVs. Due to reduced resolution after trimming the longer V34 region to shorter V4 region, the number of ASVs for cloacal, oral and tank water samples was expectedly lower (7,725 in V4 vs. 11,105 in V34 sequences).
Carapace samples had the highest median species richness and diversity (OF median 732, IQR = 420, Faith’s PD median = 45.7, IQR = 25.2; median Shannon’s index = 5.94, IQR = 2.22), which was followed by tank water, oral and cloacal samples (Fig. 5a, 5b). The differences between sample sites’ alpha diversity were detected as statistically significant (Kruskal-Wallis rank sum test), however pairwise comparisons showed differences only in Faith’s phylogenetic diversity and OF: between cloaca and all other sample sites (except tank water based on OF), oral cavity and carapace. All beta diversity metrics showed significant differences between body sites for Bray-Curtis (R2 = 0.13, Pr(> F) = 0.001), Jaccard (R2 = 0.10, Pr(> F) = 0.001), unweighted and weighted UniFrac (R2 = 0.16, Pr(> F) = 0.001; R2 = 0.18, Pr(> F) = 0.001, respectively), and robust Aitchison distances (R2 = 0.09, Pr(> F) = 0.001). Pairwise PERMANOVA detected differences between all sample site pairs (Pr(> F) < 0.01). Highly ranked ASVs impacting the distribution of samples on rPCA biplot belonged to Pseudoalteromonas sp., Vibrio sp., Shewanella sp., uncultured Saccharospirillaceae, uncultured Marinifilum and uncultured Cardiobacteriaceae (Fig. 5c). The taxonomic composition of endozoic samples and tank water using the V4 sequences resembled the one detected with V34 and it is reported in Table S5. Additional information on taxonomic composition of epizoic bacterial communities in carapace samples is reported in Supplementary Results.
Differential abundance analysis on ASVs (structural zeros excluded) detected 17 significantly DA ASVs across all sample sites (Fig. 6). A member of Rhodobacteraceae family (v4ASV8861) and Halioglobus sp. (v4ASV11062) were consistently DA in oral samples while the same goes for Cardiobacterium sp. (v4ASV10814) and a member of Enterobacteriaceae family (v4ASV11383) in cloaca. In carapace samples a member of Flavobacteriaceae family (v4ASV2446), cyanobacteria Leptolyngbya sp. (v4ASV4472), Ahrensia sp. (v4ASV8480), Sphingomonadaceae (v4ASV9910), Erythrobacter sp. (v4ASV9994), Gammaproteobacteria (v4ASV10111), Alteromonas sp. (v4ASV10327), Arenicella sp. (v4ASV10634), Psychrobacter sp. (v4ASV12017) and Vibrio sp. (v4ASV12307) were DA relative to other sample sites, while Marinomonas sp. (v4ASV11704) was DA abundant in carapace tested against endozoic samples, but not tank water. In tank water a member of Flavobacteriaceae (v4ASV2446) and NS3a marine group (v4ASV2817) were consistently DA (Fig. 6). Differential abundance analysis on taxa collapsed to species level is reported in Supplementary Results and Fig. S2.
{kind=link}