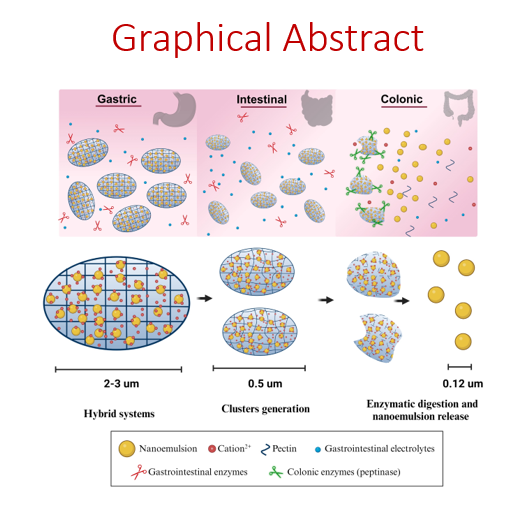
3.1. Formulation
The interaction of low methylation (< 50%) pectins with cationic species (e. g. divalent ions or polyelectrolytes) leads to the formation of pectinate gels with poor solubility and low swelling capacity in aqueous media [6, 13, 22]. Additionally, DA pectin can also affect the properties of the structure, as it increases the hydrophobic character of the carbohydrate while it facilitates the enzyme-triggered digestion of the carbohydrate backbone by the secreted enzymes from the microbiota at the colon [23, 24]. Bearing this in mind we have formulated two main PµHS families based on the condensation method of the carbohydrate (Fig. 1). Both condensation methods have been carried out with three different pectins. The three of them had similar methylation degree (around 30%) but increasing DA, i.e. 0% (NA), 13% (LA) and 20% (HA). The first attempt has been focused on ionotropic gelation using divalent cations while the second approach has been based on polyelectrolyte condensation. In both cases, the NE used for the formulation of PµHS had size values of 125 ± 5 nm, polydispersity index of 0.1 and zeta potential of – 52 ± 3 mV.
3.1.1. Ionotropic gelation
The ionotropic gelation method relies on the formation of structures due to the interaction of ionic polymers with counter ions by cross-linking. The formulation of PµHS in this work has been achieved by a modification of the ionotropic gelation method, where divalent cations, apart from the condensation of the carbohydrate backbone, have an additional role. According to the DLVO theory, ions lead to the aggregation of colloids by decreasing their electrostatic potential [15, 25], this was applied to the PµHS formulation to promote the entrapment of the nanostructure within the mesh. This aggregation induced gelation method has been carefully designed to favour i) the controlled micro-aggregation of the entrapped NE and ii) the compact packaging of these micro-aggregates into pectin microgels.
The formulation of PµHS was achieved by using three different cations, i.e. Ca2+, Ba2+ and Zn2+, that condensate pectin through its -COO− groups. As shown in Table 1 the use of pectin concentrations of 0.2% w/v with 2 mM of salt and an incubation time of 24 h led to the formation of PµHS with a mean size below 6 µm and a slightly negative ζ-potential (ranged from approximately − 12 mV to -20 mV). Shorter incubation times, lower pectin or salt concentrations resulted in the formation of non-homogeneous hydrogels with unacceptable reproducibility (data not shown). As stated before, it is believed that the NE may have a role on the condensation of the hydrogel within the microstructure that may condition its behaviour along the GIT; then pectin-microgels without NE (PµG) have been formulated and their properties included in Table 1.
The ionotropic gelation method involves the condensation of the polymer by a counterion, in this case pectin and Ca2+, Ba2+ or Zn2+, but one important issue to consider is that this interaction may be affected by the nature of the cation itself. Both, Ca2+ and Ba2+, interact by an ionic bond with the oxygen atoms of the -COO− groups of pectin, while the presence of empty d orbitals in Zn2+ favours the interaction through a coordination bond between Zn2+ and the same -O− atoms of the polysaccharide backbone [6, 22]. This was considered when designing the formulation screening of PµHS, observing that despite having the same interaction with the -COO− groups of pectin, differences could be observed in the resulting PµHS properties.
Interestingly, although both Ca2+and Ba2+ led to the aggregation-induced gelation of the components through ionic interactions, the size of PµHS formulated with Ca2+ was remarkably lower (< 1 µm) compared to the systems obtained with Ba2+ (~ 5 µm). It is reasonable to consider that a certain size of PµHS may be required to achieve a satisfactory loading of the NE, so Ca2+ PµHS would not be adequate for an efficient NE entrapment due to their low size. This was supported by the size graph of PµHS formulated with Ca2+, which clearly evidences a bimodal distribution that could be assigned to the non-entrapped NE and the PµHS (Fig. S1). These results led us to exclude this formulation for the rest of the experiments. Opposite, the higher size values achieved when Ba2+ was used as aggregation-induced gelation agent may ensure a better entrapment of the NE in the microgel structure. This difference in size could be explained by the bigger atomic mass of Ba2+, compared to Ca2+, that might facilitate the aggregation and the entrapment of NE in the microgel structure.
On the other hand, PµHS formulated with Zn2+ had size values between those of Ca2+ and Ba2+ PµHS. In this sense the aggregation induced gelation of the formulation through a coordination bond between Zn2+ and -COO- groups resulted in an adequate size to ensure the entrapment of the NE droplets but also facilitate its enzyme-triggered release at the colon.
The PµG systems, pectin-microgels without NE, formulated through the ionic binding of either Ca2+ or Ba2+ with pectin have heterogenous and non-reproducible size distributions (Table 1). Despite the well-known properties of Ca2+ and Ba2+ to condensate pectin and other polysaccharides as hydrogels [6], it seems that, under these formulation conditions, the presence of the NE as a condensation core is required to favour the assembling of the microstructure [11]. The use of Zn2+ as a condensation agent led to microgels of 2.5–3.5 µm and low size dispersion. In this context, we use the PµG formulated with Zn2+ to understand the role that NE have within the structure of the PµHS along the GIT.
Table 1. Physicochemical properties of PmHS obtained by aggregation induced gelation
|
PmHS
|
PmG
|
|
Pectin
|
Cation
|
Size (µm)
|
z-Pot (mV)
|
Size (µm)
|
z-Pot (mV)
|
|
NA
|
Ca2+
|
0.7 ± 0.3
|
-18.7 ± 0.8
|
Non-reproducible
|
|
Ba2+
|
5.1 ± 3.6
|
-20.1 ± 5.1
|
Non-reproducible
|
|
Zn2+
|
2.6 ± 0.3
|
-12.5 ± 0.4
|
2.4 ± 0.1
|
-13.0 ± 0.4
|
|
LA
|
Ca2+
|
0.4 ± 0.1
|
-18.2 ± 1.2
|
Non-reproducible
|
|
Ba2+
|
5.3 ± 3.5
|
-14.8 ± 0.4
|
Non-reproducible
|
|
Zn2+
|
2.3 ± 0.4
|
-12.1 ± 0.3
|
3.5 ± 0.2
|
-11.6 ± 0.1
|
|
HA
|
Ca2+
|
0.6 ± 0.3
|
-17.1 ± 4.4
|
Non-reproducible
|
|
Ba2+
|
5.6 ± 3.5
|
-13.7 ± 2.0
|
Non-reproducible
|
|
Zn2+
|
1.5 ± 0.1
|
-14.1 ± 0.5
|
3.4 ± 0.1
|
-14.1 ± 0.5
|
3.1.2. Polyelectrolyte mediated condensation
As alternative to the ionotropic gelation, we also formulated PµHS through a polyelectrolyte mediated condensation. For that purpose, the negatively charged NE were modified by its coating with positively charged BSA (isoelectric point ranging from 5.1–5.5) [19, 26, 27], in order to obtain a positive net charge of the system able to interact with the anionic moieties of the polysaccharide. The protein adsorption onto the NE surface was promoted by adjusting the pH of the BSA just below its isoelectric point [19, 28]. As shown in Table 2, the three coating densities tested (i.e. 4.5, 9.0 and 13.5 mg/m2) led to the charge inversion of the NE, from the negative values of − 52 mV of the uncoated NE. However, those NE formulated with the lowest coating density had more heterogeneous size and ζ-potential distributions. Coating densities equal or higher than 9.0 mg/m2 resulted in formulations with similar size and superficial properties, suggesting the saturation of the NE surface once coating densities equal or higher than 9.0 mg/m2 were achieved [26, 27].
The PµHS obtained by the condensation of the different pectins with the BSA-coated NE had size values below 10 µm and negative superficial charge. The increase of size and charge inversion observed clearly showed the efficacy of this method to entrap the cationized NE in the pectin microgel for all the tested pectins. The size values of these systems were considered adequate as they guarantee the incorporation of the NE to the PµHS and may also facilitate the digestion of the structure by the enzymes secreted by the microbiota at the colon.
Unfortunately, the PµHS prepared with an inner BSA coating density of 4.5 mg/m2 had poor colloidal stability after the formulation (< 24 h). These results can be attributed to non-dense enough BSA coatings of the NE, compromising the properties of the BSA-coated NE as condensation cores for the pectin [27]. Instead, the PµHS formulated either at BSA coating densities of 9.0 or 13.5 mg/m2 had acceptable colloidal stability after formulation. The similar properties of the PµHS formulated either at 9.0 or 13.5 mg/m2 are in line with the physicochemical properties observed for both BSA-coated NE and suggest a saturation of the NE surface at 9.0 mg/m2 [19]. Bearing this in mind the prototypes formulated at a BSA coating density of 9.0 mg/m2 were selected for further studies.
Table 2. Physicochemical properties of PmHS obtained by polyelectrolyte condensation
|
|
BSA-coated NE
|
|
|
PmHS
|
|
|
BSA (mg/m2)
|
Size (µm)
|
z-Pot (mV)
|
Pectin
|
Size (µm)
|
z-Pot (mV)
|
|
4.5
|
0.7 ± 0.5
|
16.5 ± 4.4
|
NA
|
7.3 ± 1.7
|
-30.1 ± 1.0
|
|
LA
|
1.9 ± 0.2
|
-30.1 ± 1.0
|
|
HA
|
3.5 ± 0.3
|
-25.3 ± 0.6
|
|
9.0
|
0.2 ± 0.1
|
19.8 ± 0.8
|
NA
|
1.7 ± 0.2
|
-33.2 ± 0.7
|
|
LA
|
1.5 ± 0.1
|
-32.0 ± 0.9
|
|
HA
|
2.1 ± 0.1
|
-31.1 ± 1.1
|
|
13.5
|
0.2 ± 0.1
|
18.6 ± 0.7
|
NA
|
1.4 ± 0.1
|
-29.8 ± 0.7
|
|
LA
|
1.4 ± 0.1
|
-30.7 ± 0.7
|
|
HA
|
2.3 ± 0.2
|
-30.1 ± 1.0
|
3.2. Stability along the GIT
The stability of PµHS along the GIT can be compromised due to its harsh conditions; therefore, we have separately analysed one by one the effect of each variable of the simulated GIT media (i.e. pH, electrolytes and enzymes) on the integrity of the systems. Undoubtedly, the type of interaction that allows the PµHS formation and the physicochemical properties of the associated pectin may determine their stability, so they were also evaluated.
3.2.1. Structural integrity in SGF
The analysis of the structural integrity of the different PµHS prototypes in simulated gastric fluid (SGF, USP XXIX) was performed for 2 h in SGFs and afterwards in SGF. As can be observed in Fig. 2, the results indicated that the condensation method as well as the physicochemical properties of pectin had a clear effect on the structural integrity of the PµHS in this media.
Ionotropic gelation.
The DA of pectin was determinant in the stability of PµHS formulated with Ba2+ as aggregation induced gelation agent. Indeed, it was observed that NA prototypes experienced a strong size reduction, meanwhile PµHS prepared with the amidated pectins (either LA or HA) displayed a clear size increment just after incubation in SGFs or SGF, the NA prototype presented a clear size reduction (x10) under the same conditions.
The structural viability of these PµHS prototypes depends on the inner electrostatic interactions of the NE, the divalent cation and -COO− groups of the pectin, but also of the inner attractive forces between the NE and hydrophobic moieties of the pectin and the repulsive electrostatic interactions that avoid the microparticle-microparticle aggregation [14, 15]. The charge cancellation of both superficial groups of the NE and -COO− groups of NA pectin may be the reason behind the disassembling of the microstructure [14]. The comparison between SGFs and SGF indicated that pepsin did not affect the structural viability of this prototype.
Contrary to the results of the NA prototypes, the amidated PµHS clearly increased their size under gastric conditions. From a pharmaceutical point of view, the DA of pectin may improve the performance of hybrid systems, as it facilitates the enzyme-triggered digestion of the hydrogel under colonic conditions [23, 24]. Interestingly, higher DA values lead to carbohydrates more hydrophobic, a relevant aspect to consider from a structural point of view. In absence of the electrostatic interactions, it seems that the hydrophobic interactions between the NE and the amidated pectins hampered the disassembling of these microstructures. Unfortunately, the cancellation of the superficial net charge of the microgel under acidic conditions may be reason behind the massive aggregation of these structures under gastric conditions [16, 29].
The DA of pectin also determined the stability of those PµHS obtained with Zn2+. The formulation with NA pectin led to the disassembling of the microstructure after 2 h of incubation either in SGFs or SGF, as observed for Ba2+. Although it has been described that Zn2+ may form stronger interactions with pectins than Ca2+ or Ba2+, it seems that the micrometric size of these PµHS and the reduced hydrophobicity of the non-amidated pectin contribute to the sensitivity of these formulations to the external stimuli [14, 25]. On the contrary, the amidated pectins improved the stability of the systems in SGFs and SGF, with a slight size increase after 2 h in gastric media.
These results suggest that using Zn2+ instead of Ba2+ as aggregation induced gelation agent may improve the structural viability of PµHS under gastric conditions [6, 8].
Polyelectrolyte mediated condensation.
As observed for Ba2+ and Zn2+, PµHS formulated with NA showed a clear size reduction after their incubation in SGFs and SGF. The low pH of the gastric fluid may increase the superficial charge of the BSA-coated NE, but it also decreases the fraction of anionic -COO− groups of the pectin backbone, minimizing the electrostatic attraction between the NE and the carbohydrate, which can compromise the structural stability of PµHS.
Those systems obtained with LA had a similar behaviour in both SGFs and SGF, showing a clear size increase along the incubation time. These results can be explained by the reduction of the superficial charge of the PµHS derived by the charge cancellation of the outer pectin shell [16, 25]. The medium composition affected the structural viability of PµHS formulated with HA pectin. The PµHS incubated in SGFs clearly aggregated, but the kinetic of the aggregation was slowed down when the pepsin was in the gastric medium. This effect may be attributed to the coating of the PµHS by pepsin, that can partially stabilize the microparticles [19, 27].
3.2.2. Structural integrity in SIF
The maintenance of the structural integrity of the PµHS prototypes in simulated intestinal fluid (SIF, USP XXIX) was performed for 4 h. The effect of pancreatin and electrolytes on the stability of the systems was studied separately by incubating them in SIFs or SIFe. Afterwards, the structural integrity in SIF medium (USP XXIX) was evaluated. The results obtained indicated that the behaviour of PµHS prototypes depended on: i) the formulation method applied, ii) the properties of the pectin used and iii) the components of the intestinal media (Fig. 3).
Ionotropic gelation.
The PµHS formulated with Ba2+ by the ionotropic gelation technique did not experience a remarkable effect on the size distribution after their incubation in SIFe, independently of the pectin used (i.e. NA, LA or HA). This behaviour could be attributed either to the proper stability of the formulation or to the formation of a homogeneous enzymatic coating onto the PµHS that promotes their stabilization [16, 19, 27, 30].
Nevertheless, the incubation of those prototypes in SIFs resulted in the disassembling of the microgels. Interestingly, the final size of PµHS was clearly dependent on the pectin used, i.e. ∼ 0.4 µm for NA, ∼ 0.3 µm for LA and ≈ 0.2 µm for HA, showing stronger disintegration those PµHS formulated with amidated pectins. This behaviour, which is contrary to the one observed under gastric conditions, can be due to the screening of the ionic bonds between the components that constitute the PµHS by the electrolytes of the media [14]. Indeed, the increase of the DA in the pectin backbone and the presence of the electrolytes in the media may reduce the availability of free carboxyl groups.
The study performed in SIF emphasized even more the differences observed in the behaviour of PµHS. Those systems formulated with NA pectin experienced the slightest variation of size along the assay, being comparable to the PµHS when incubated in SIFe. Meanwhile, PµHS formulated with HA displayed a similar behaviour that in the presence of SIFs, with final size values of ∼ 0.2 µm; the PµHS prepared with LA had an intermediate response, with final size values of ≈ 0.4–0.5 µm, which is remarkable higher than the observed in SIFs.
These results indicated that the DA of pectin may determine the interaction of Ba2+ constituted PµHS with the media. The use of NA pectin may favour the interaction of the microgel with the surrounding pancreatin to quickly form a protein corona able to stabilize the structure and avoid a fast disassembling of the microgel by the SIF electrolytes [27]. Nevertheless, the formation of the stabilizing protein corona was not favoured when HA constituted the PµHS, leading to the disruption of the structure by the electrolytes of SIF [26].
Curiously, DA of pectin did not remarkably affect the integrity of PµHS formulated with Zn2+ in SIF. All the prototypes incubated in media including pancreatin (SIFe) experienced a slight reduction of their mean size, meanwhile the presence of electrolytes clearly promoted their disassembling (SIFs). Indeed, once PµHS formulated with Zn2+ were incubated in SIF, the final size was quite similar to the one obtained just with electrolytes. Nevertheless, it is important to highlight that despite the disassembling of Zn2+ PµHS, their final size values were remarkably larger (≈ 0.3–0.4 µm) than those of bare NE (∼ 0.1 µm). This result may indicate the formation of small clusters of NE droplets still associated to pectin after the incubation of PµHS in SIF.
Polyelectrolyte mediated condensation.
The incubation of the prototypes obtained by the polyelectrolyte mediated condensation technique in SIF led to the same results regardless of the medium composition or the DA of the pectin. All the PµHS prototypes had size values of ∼ 0.2–0.3 µm after their incubation, which were like those of the original BSA-NE cores (0.2 ±0.1 µm) used to formulate PµHS. To shed some light on these results, we analysed the average mean size of the prototypes formulated with the three pectins in phosphate buffer (pH 7) of low ionic strength (2 mM). Interestingly, all the microgels presented the same behaviour as can be observed in Fig. 3c. These results indicated that the neutral pH of the intestinal milieu is enough to promote the total disassembling of the PµHS. Polyelectrolyte condensation have been widely used in the pharmaceutical research field to formulate nanoparticles and microparticles for different medical needs [2, 6, 8, 13]. The rationale behind the design of these structures relies on the interaction of two oppositely charged polyelectrolytes. The adsorption of BSA onto the bare NE was performed at pH just below the protein isoelectric point to promote both electrostatic and hydrophobic interactions with the NE [19, 31]. Therefore, the BSA shell formed on the NE gives positive net charge and a hydrophilic character to the BSA-NE complex [19, 26]. Bearing this in mind, the incubation of the PµHS at pH ≈ 7 led to the charge inversion of the BSA shell (isoelectric point ranging from 5.1–5.5) while it maintained its hydrophilicity. Under this scenario, at pH ∼7 none of the pectins can bind to the BSA-NE either by electrostatic or by hydrophobic interactions. These results led us to exclude the PµHS prototypes obtained by the polyelectrolyte mediated condensation technique for the subsequent experiments.
3.2.3. Structural integrity in SCM
The structural integrity of the PµHS prototypes obtained with Ba2+ or Zn2+ in simulated colon media (SCM) was determined by independently evaluating the effect of the media components. Once the medium was buffered with HEPES, the first studies were performed by adding exclusively pectinase to the medium (SCMe). Afterwards, the effect of the electrolytes was determined (SCMs) to finally evaluate the behaviour of the systems in complete SCM [10], see Fig. 4.
The PµHS formulated either with Ba2+ or Zn2+ experienced a biphasic response in the presence of enzymes, which was more acute for the Ba2+ prototypes. The biphasic profile had an initial phase of size reduction of the PµHS followed by a size growth phase, that could be attributed to the avidity of pectinase for the pectin outer shell of the system. The pectinase-PµHS interaction may result in an initial size reduction based on the fast assembly of pectinase coated clusters of microgel followed by a degradation phase. Under this scenario, the most feasible hypothesis is the creation of a protein corona that fixes the size of the microgel as it degrades the carbohydrate matrix [20, 25, 30, 31].
As can be seen in Fig. 4, the electrolytes of SCM favoured the disassembling of the PµHS in clusters of ≈ 0.2–0.4 µm. The response to the electrolytes obtained for the Zn2+ PµHS was more progressive than the one observed for the Ba2+ prototypes. Indeed, this behaviour is similar to that achieved in SIF and can be explained by the screening of the NE-cation2+-pectin due to the electrolytes [14]. Therefore, Zn2+ PµHS were selected for the following studies.
Finally, the combination of the enzyme and the electrolytes led to a similar biphasic response to that observed just in the presence of pectinase (SCMe). In this case, the explanation could be that the presence of the electrolytes could facilitate the digestion activity of pectinase, ending up in a faster release of the NE droplets from the microgel. However, to confirm this hypothesis it is mandatory to carefully analyse the dynamics of the NE entrapped in the pectinase-PµHS clusters under colonic conditions.
3.2.4. Role of the NE on the structural integrity of PµHS
The gelation induced aggregation method relies on the formation of pectin-based hybrid systems in the micrometric range by a combined effect of aggregation of the NE by the presence of cations, together with the cross-linking of the carbohydrate by divalent cations. Within this context, it would be required to understand the role of the NE in the PµHS stability; therefore, the size evolution of PµG formulated with Zn2+ under GIT conditions was evaluated. In all the tested media, the three PµG prototypes were totally unstable in the presence of electrolytes, resulting in a quite fast size increase. This is in line with our hypothesis about a disruption of the microgel matrix when electrolytes screen the binding interactions that condensate these microgels [14]. The incubation of PµG in SGF and SIF in the presence of enzymes led to a similar response that of PµHS. The surfactant character of the GIT enzymes will ensure that, once in contact with the microgel, they will adsorb onto its surface. Pepsin and pancreatin are not able to degrade the carbohydrate matrix of neither PµHS nor PµG, but the formation of the protein corona will fix the size of the microgel [25, 30, 31].
Interestingly, the behaviour of PµG in SCMe and SCM was clearly different to the one observed for the PµHS prototypes, that had an initial size reduction followed by a slight size increase along the next 24 h. In this case, the incorporation of the pectinase to the medium did not end up on the formation of a protein corona over the PµG prototypes but led to an initial size reduction of the microgel followed by a fast disintegration of the microgel particles.
The differences found in the behaviour of PµHS and PµG under SCM conditions suggest that the entrapped NE are clearly conditioning the enzyme-triggered degradation of the pectin microgel matrix. These results evidence that the presence of the NE in the PµHS may play a structural role that helps to maintain the hydrogel structure during enzymatic degradation [11]. To get a better insight on the behaviour of the entrapped NE under colonic conditions we have evaluated by particle tracking the dynamics of the NE entrapped in PµHS formulated by the aggregation induced gelation with Zn2+.
3.3. Enzyme triggered release: Dynamics of NE in the PµHS
From a classical point of view, the physicochemical characterization of nano- and micro-medicines for oral delivery has been focused on the response of the size and ζ-potential of the studied prototypes as a function of the medium composition. However, for hybrid systems it is mandatory to understand not only the structural modifications of the formulation, but also the dynamics of the entrapped nanostructure as a function of the external stimuli. Under this scenario, particle tracking has arisen as an excellent tool to analyse the behaviour of each single particle as a function of the external stimuli [11, 12]. Focused on the PµHS, particle tracking may shed light on the variables involved on the success of the enzyme-triggered release of the NE from the carbohydrate matrix at the colon.
The approach followed for the evaluation of the dynamics of the NE in the PµHS was to determine the implications that each individual stimuli had on the response of the prototypes to colonic conditions. In the absence of electrolytes or enzymes, NE presented a Dc of 1.8*10− 3 µm2/sec. The ratio between the Dc of the entrapped NE with respect the free NE clearly demonstrated that the NE were firmly retained inside the PµHS after their formulation (Dc/DcNE = 4.5*10− 4) [17, 21, 30]. This result supports that the formulation of PµHS by the aggregation induced gelation method allows the tight entrapment of NE inside the microstructure.
The incorporation of pectinase or electrolytes into the medium increased the Dc/DcNE values by two orders of magnitude (Fig. 5), indicating that both components intensely affect the dynamics of the NE entrapped inside the carbohydrate hydrogel. This dramatic increase in the diffusion of the NE within the hydrogel may be considered as the first stage for the following release of the oil droplets from PµHS. Among both components, the most intense effect on the diffusion of the NE was observed in SCMs (Dc/DcNE ∼ 0.1–0.17), while the presence of pectinase slightly decreased the response of the PµHS (Dc/DcNE ∼ 0.04–0.09). This is in line with our structural stability results, where the electrolytes usually affect more aggressively PµHS and PµG structures. Interestingly, after the initial increase, the Dc/DcNE values remained barely the same throughout the incubation time (up to 24 h). These results suggest that the incorporation of the NE to the microgel matrix may help to preserve to some extent the structural integrity of the hydrogel matrix even in the presence of pectinase. This structural role of the NE in PµHS may also determine the release of the NE themselves, by controlling its diffusion inside the carbohydrate matrix over time [11, 12].
Particle tracking is a powerful technique that not only provides information about the mean or ensemble diffusion coefficient, but it also facilitates information about the diffusion mode of each individual particle [17, 20]. The value of the anomalous diffusion parameter (α) depends on the diffusion pattern of the particles, having free-diffusing particles an α ≈ 1, while those nanoparticles with a hampered diffusion present α < 1. Depending on the NE entrapment, the transport mode of each individual oil droplet can be sorted as follows: (i) immobile (α < 0.2); (ii) hindered (0.2 < α < 0.4); (iii) subdiffusive (0.4 < α < 0.9); and (iv) diffusive (0.9 < α) [17, 20, 21]. Bearing this in mind, we calculated the α values of each individual oil droplet of the NE entrapped in PµHS when incubated in buffer solution, in the presence of enzymes (SCMe), electrolytes (SCMs) or both together (SCM). The results obtained for the PµHS formulated with LA pectin are included in Fig. 6, but similar results were obtained with NA and HA pectin.
The incubation of PµHS in low ionic strength buffer solution indicated that more than 70% of the oil droplets were totally stuck inside the carbohydrate matrix, with an approximately 10% of population as hindered and subdiffusive droplets. It is interesting to note that around a 3% of the NE droplets displayed a free diffusion behaviour. This small population may correspond to those weakly bounded oil droplets placed in the outer shell of the PµHS. Altogether, these results support that the aggregation induced gelation method is an efficient technique to incorporate nanostructures into micrometric hybrid systems.
The presence of enzymes or electrolytes in the media, separately or combined, led to an immediate modification of the diffusion mode of the entrapped NE. As shown in Fig. 6, these elements remarkedly reduced the population of immobile oil droplets from ≈ 70% down to ≈ 15%, while the hindered fraction displayed a slight increase. Additionally, subdiffusive and diffusive fractions experienced a 4-fold and 6-fold increase respectively. Despite the differences showed in the ensemble Dc/DcNE when electrolytes, enzymes, or the combination of both elements were present (Fig. 5), α distribution values displayed a similar pattern independently on the stimuli. The NE entrapped in NA or HA PµHS displayed a similar distribution of the diffusion mode as LA (data not shown).
As observed for the ensemble diffusion coefficient (Fig. 5), the distribution of the α values was almost constant along the incubation time, while the release of the NE from the carbohydrate matrix led to a reduction in the number of available trajectories over the time (Fig. S2). These results suggest that electrolytes and enzymes promote a steady state that controls the release of NE from the matrix of hybrid systems. Under this scenario, more than 60% of the remaining NE droplets displayed an active diffusion regime (α > 0.4) along the incubation time (data not shown).
Based on the behaviour of the ensemble Dc and α as a function of medium composition, pectin and incubation time, we evaluated the distribution of the Dc of each individual oil droplet in SCM. Figure 7 shows the distribution of Dc values of the NE entrapped in the different PµHS in buffer or SCM just after incubation and at 24 h. The cumulative fraction of oil droplets was sorted as follows: (1*10− 4)-(1*10− 3); (1*10− 3)-(1*10− 2); (1*10− 2)-(1*10− 1) and (1*10− 1)-1 µm2/sec. As expected from the α values, most of the NE droplets of the LA PµHS were highly retained in the carbohydrate hydrogel when incubated in the low ionic strength buffer, showing more than 80% of the oil droplets a Dc < 1*10− 2 µm2/sec. Similar results were obtained with the others pectin (data not shown).
The incubation of the systems in SCM led to an instant change in the Dc profiles of the NE (Fig. 7a). All the PµHS formulations had a minimal fraction of NE with Dc < 1*10− 2 µm2/sec (< 5%). Interestingly, the cumulative fraction of NE droplets at higher Dc was affected by the used pectin as follows:
-
1*10 − 2 < Dc < 1*10− 1: The cumulative fraction of NE droplets was proportional to the DA of the used pectin.
-
1*10 − 1 < D c : The cumulative fraction of the NE droplets was higher than the observed in the previous range and inversely proportional to the DA of the pectin.
As shown in Fig. 7b, the NE entrapped in the different PµHS behaved as follows after 24 h of incubation in SCM:
-
D c < 1*10− 2: All the PµHS presented a slight increase in the cumulative fraction of oil droplets with respect to the initial times of incubation (Fig. 7a).
-
1*10 − 2 < Dc < 1*10− 1: The cumulative fraction of NE droplets increased with DA. The population of the NE droplets from all the PµHS displayed a reduction with respect to the values observed just at the beginning of the incubation (Fig. 7a).
-
1*10 − 1 < D c : Those PµHS formulated with amidated pectins displayed a reduction on the cumulative fraction of oil droplets with respect to the previous Dc range, meanwhile the use of NA pectin increased the population of NE droplets with respect to the previous Dc range. In any case, all the formulations presented a reduction of the cumulative fraction of NE droplets with respect to the initial incubation time (Fig. 7a).
The particle tracking studies clearly demonstrated that during the enzyme triggered-release of PµHS, the dynamics of the NE depend not only on the presence of enzymes, but also on the electrolytes and the incubation time. In addition, it can be concluded that the initial incubation times favoured the presence of populations with a higher Dc value than longer incubation times. To understand the shift of the NE population towards lower Dc values it is necessary to consider the number of available trajectories as a function of the time. As commented above, due to the release of the NE from the carbohydrate matrix, the number of available trajectories decreases as the incubation time increases. Under this scenario, it seems that the PµHS are coated by the pectinase and their carbohydrate matrix is degraded by a surface-core process, where the remaining population of NE correspond to the less available oil droplets [16, 30].
Additionally, it has been described in the literature that high DA favours the degradation of pectin backbone by pectinases [6, 23, 24]. As mentioned above, the DA increases the hydrophobicity of the carbohydrate [6], this may have a structural role on the formation of the PµHS affecting the response of the systems to the enzymes. During the aggregation induced gelation process the incorporation of NE into the PµHS will face these hydrophobic residues towards the surface of the oil droplets, increasing the overall structural strength thanks to the attractive Van der Waals interactions [18, 19, 27]. Within this context, and taking into consideration the particle tracking results, it is feasible to hypothesize that the hydrophobic residues of pectin will not be totally accessible to the enzymes for the enzymatic degradation of the carbohydrate matrix. This structural role of the DA may explain the unexpected response of PµHS to the enzyme-triggered release of the NE from the carbohydrate matrix.
As observed from the structural stability and the particle tracking results, it is worth to highlight the relevant role of electrolytes on the behaviour of the PµHS. Our results show that the pectinase-triggered release of the NE under colonic conditions is not only favoured by the enzyme itself, but electrolytes may play a relevant role on the performance of hybrid systems.
The formulations aimed for colonic drug delivery can be administered by the oral route or via the rectum, having both approaches either advantages and limitations, mostly related to patients’ compliance, poor absorption, or instability due to the harsh conditions of the GIT. In this context, the proper design of the properties of formulations is decisive for their performance. Indeed, the evidence that smaller size values may improve the performance of the formulations at the colon prompted us to design hybrid systems at the micrometric range [3–5]. Considering that these systems are aimed for the administration by the oral route, the behaviour of the entrapped NE under colonic conditions may be also affected by the electrolyte-mediated partial disassembling of the PµHS observed in SIF. Then, the last set of experiments studied the effect that the preincubation of PµHS in SIF may have on the dynamics and release of the entrapped NE under colonic conditions.
The intermediate behaviour shown by the LA pectin compared to the other pectins in the previous studies led us to select the PµHS formulated with this pectin for the following experiments. The incubation of the PµHS in SIF for 4 h at 37 ºC was followed by an additional one in SCM, where the diffusion of the entrapped NE was determined at 10 min, 3 h, 8 h and 24 h by particle tracking. These systems were compared with a control group of PµHS directly incubated in SCM, and the ensemble Dc of the NE from both groups were plotted in Fig. 8.
The preincubation of the PµHS in SIF had two main effects on their behaviour under simulated colonic conditions. Firstly, the partial disassembling of the PµHS in SIF significantly improved the enzyme-triggered release of the NE at the early stages of the incubation in SCM (higher ensemble Dc values). Secondly, the preincubation in SIF resulted in systems with smaller size values but more dispersed. This size heterogeneity clearly affected to the following enzyme-triggered release of the NE at the initial incubation times in SCM (less homogeneous Dc values distribution).
Finally, as the digestion of the PµHS was progressing, the Dc evolved towards more homogenous but lower values. As commented above, only those oil droplets that have a more restrictive access to pectinase will remain available for long incubation times in SCM, leading to the homogenization and reduction of the ensemble Dc.
A more detailed analysis of the Dc distribution showed that the preincubation of PµHS in SIF clearly shifted the NE population towards higher Dc values (Fig. 9). This modification of the NE dynamics indicated that the preincubation of PµHS in SIF facilitated the enzyme-triggered release of the NE under simulated colonic conditions. More interestingly, the results demonstrated that the preincubation in SIF improved the Dc distribution not only at the initial stages of the incubation in SCM but also after 24 h of digestion. From a quantitative point of view, the preincubation of PµHS in SIF produced an increment of the cumulative fraction of the fastest NE droplets of ≈ 10% and ≈ 25% at the initial and final times of incubation respectively.
Overall, particle tracking was crucial to understand the effect that enzymes and electrolytes have in the dynamics of a nanostructure entrapped in the PµHS. In this sense, we observed that both stimuli determine the dynamics of the NE in PµHS, which can be then identified as a multi-stimuli formulation for colonic delivery via oral route. This is an added advantage, since relying on a single physiological stimulus (e.g. time, pH or enzymatic digestion) can lead to great variability, as physiological modifications would hinder drug release [1]. Considering the pathophysiological process that occur in ulcerative colitis it is reasonable to propose a multi-stimuli system for colonic drug delivery, in this case able to relax the hydrogel matrix due to the effect of the intestinal electrolytes, for the subsequent enzymatic activity of the pectinases produced by colon microbiome. Additionally, the particle tracking results have shown that the effect of the DA on the digestion of the PµHS at the colon may be masked by the presence of the NE. Our results suggest that, under these conditions, the main role of DA is to increase the overall structural strength of the PµHS rather than facilitate the action of pectinases.
These results have allowed the identification of a multi-stimuli formulation based on PµHS that can be applied as a versatile platform to the required medical need. The possibility of using antioxidant components, such as α-tocopherol or ascorbyl derivatives, for formulating NE could be of great interest when oxidative processes are involved in the pathophysiology of the disorder, like in ulcerative colitis. In addition, the capacity of NE to encapsulate different active molecules allows to adapt the formulation to the specific requirements of the disease. All in all, these experiments have described the conception of a salt and enzyme-mediated multi-stimuli system for colon-targeted, which potential in drug delivery will be elucidated in future studies.
{kind=link}