A number of researches have demonstrated the ability of bacterial vesicles to fuse with eukaryotic membranes, facilitating the penetration of OMVs contents into the cell [26–28]. The overall biological effects observed in OMV-treated cultures may contribute to discovering new mechanisms of host-microbiota interaction. However, preliminary proteomic and metabolomic characterization of bacterial vesicles may be useful for detailed functional annotation of molecular cellular effects treated with OMVs. A previous study dedicated to proteome and metabolome annotation of Bacteroides fragilis OMVs, was continued by studying the proteomic effects of OMVs treated cell lines [9].
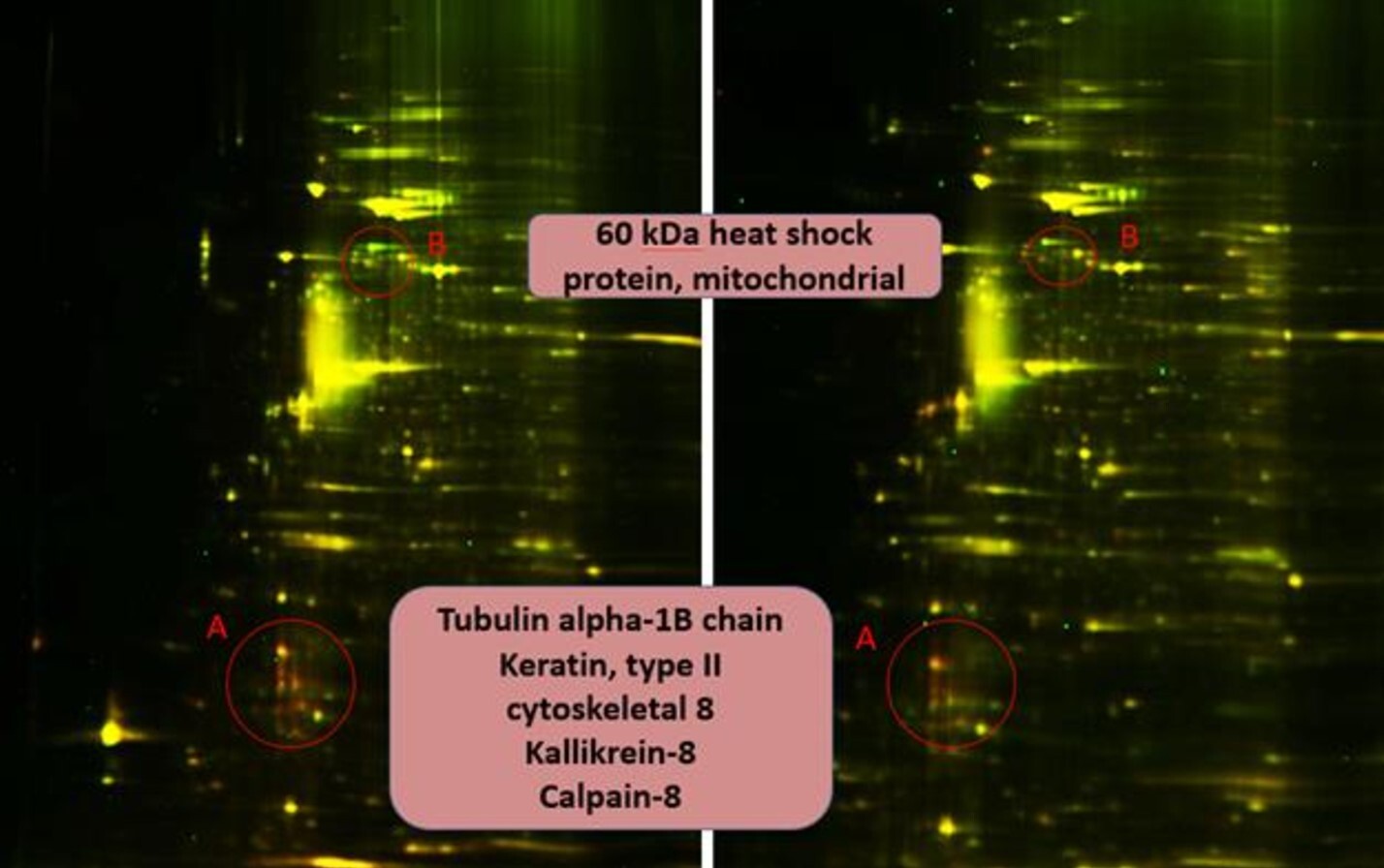
At the beginning the quality and concentration of the isolated vesicles were assessed by using TEM and NTA methods. We confirmed that morphology and concentration of isolated OMV were in complete agreement with both literature data and our previously published data. [21]. As ETBF has previously been shown to secrete a vesicle-associated toxin, the SW837 cell culture was treated with ETBF OMVs [29]. As a result, the time-dependent E-cadherin degradation was also confirmed in the SW837 cell line, as previously observed in the HT-29 cell line [25]. Despite the fact that OMVs containing toxin contributed to the E-cadherin degradation, vesicles had no effect on the morphology and viability of the cells. It is important to note that isolated toxin contributes to the reversible cell rounding and separation [10, 11]. Perhaps the destructive effects of the toxin to the whole cellular physiology are compensated by the other proteins' action. Previously, the secretome of the HT-29 cell lines treated with the isolated Bacteroides fragilis toxin has been fully described [25]. The protein numbers which were dysregulated in the cell line treated with the toxin were minimal compared to the untreated cell line and amounted to approximately 20–30 proteins. ETBF OMVs contain more than 300 proteins including enzymes and we have expected a large number of molecular effects in HT-29 cell lines treated with ETBF OMVs. However, the same numbers of proteins were found up and down regulated in HT-29 cell lines treated with ETBF or NTBF OMVs for 5 hours. We found that proteins belonging to the cell cytoskeleton (tubulin alpha-1B chain, keratin type II cytoskeletal 8, kallikrein 8, calpain-8) were down regulated, but heat shock proteins - chaperones were up regulated in both HT-29 cell lines treated with ETBF and NTBF OMVs. The identified mitochondrial heat shock protein (60 kDa) is responsible for the folding of the new proteins entering to the cells and may be involved in the processes of recognition and subsequent proteasomal degradation of newly arrived bacterial proteins [30]. Thus, the major differences were observed only in a group of cytoskeletal proteins. No direct effect of the toxin associated with ETBF OMV on the proteome of HT-29 cell lines was detected. The obtained result probably means that HT-29 cell lines may not be fully accessible for bacterial vesicles due to the surface mucin layer [31].
The SW-837 cell line was treated with ETBF and NTBF OMVs for 3 and 5 hours. We chose this time period for our experiment because we believed that OMVs - intestinal cell interaction time coincides with the time of normal intestinal motility [32]. Therefore, we expected to detect the proteomic effects in cell culture treated with OMVs during 3 hours and to continue to observe described effects after 5 hours of OMVs exposure.
It was possible to identify up and down regulated proteins in SW-837 cells treated with NTBF OMVs for 3 hours, which were characteristic for cellular responses from the cell differentiation to the cell’s apoptosis. Among identified proteins were: cytoskeletal components and structural proteins of the Golgi apparatus, cell adhesion proteins, proteins responsible for proteasomal degradation and enzymes active in processes of cell proliferation and transformation. According to the data obtained, a cytoskeleton reorganization has taken place for the first three hours in cultures treated with ETBF or NTBF OMVs. If some cytoskeletal proteins can be involved in the process of cellular uptake of bacterial vesicles, then other proteins such as ACTG could be immediately activated to promote cell proliferation or apoptosis [33]. Upregulated heat shock proteins are also involved in cytoskeletal reorganization [34]. Despite a number of proteomic changes, only a few groups of proteins formed significant pathways which were activated in cell lines treated with ETBF and NTBF OMVs for 3 hours. Cell-extracellular matrix interactions (HSA-446353) and Protein processing in endoplasmic reticulum (hsa04141) signaling pathways were activated in cell lines treated with NTBF or ETBF OMVs respectively. Proteins that are activated in the HSA-446353 pathway are directly involved in the reorganization of the cell cytoskeleton, probably due to the cells and vesicles membrane fusion.
However, it is also interesting to observe up and down regulated proteins that were different in cultures treated with ETBF or NTBF OMVs for 3 hours and were not associated with signaling pathways. The proteins SNN, SPB5, DTX and ARPC2 were upregulated in the SW837 cell line treated with NTBF OMVs for 3 hours. SERPINB5 is involved in cell matrix organization, regulation of epithelial cell proliferation and blocks tumor growth, invasion and metastatic process [35]. Increased SERPINB5 expression was associated with a worse prognosis in patients with colorectal cancer [36]. Upregulated SERPINB5 which was founded in culture treated with NTBF OMVs may indicate the intensification of cell proliferation, which generally can be a positive vital effect for cells. The ARPC2 protein which forms the basis of the Arp2/3 protein complex is involved in the process of actin polymerization in cells [37–38]. To date, there is no clear link that has been established between this protein and colorectal cancer, but there is evidence that ARPC2 protein was found to be overexpressed in gastric cancer. ARPC2 protein is also involved in cell proliferation and invasion [39]. Thus, like SERPINB5, the ARPC2 protein may promote cell survival when it’s treated with the NTBF OMVs. DTX4 was also upregulated in cell cultures treated with NTBF OMVs. DTX is a ubiquitin ligase and promotes the degradation of the TBK1 protein, a participant in the interferon-I mediated inflammatory pathway [40]. The inflammatory process is initiated by innate immune cells with the following activation such pathways as the NF-κB, MAPK and type I interferon (IFN-I) mediated inflammatory pathway. Pattern recognition receptors have their intracellular representation as NLR receptors, genetic mutations in which, for example, in the NOD2 gene are associated with susceptibility to Crohn's disease [41]. NLRs are known to have indirect roles of NF-κB regulation, MAPK-pathway activation with the following secretion of cytokines, chemokines and IFN. DTX4 protein found to be overexpressed in cell lines treated with NTBF OMVs may contribute more active ubiquitin associated degradation of TBK1 leading to a decrease activity of the participants of interferon-mediated inflammatory pathway. Interestingly, Bacteroides fragilis vesicles reduce the inflammatory response by PSA mediated immune suppression [42]. It is likely that the overall anti-inflammatory effect of vesicles may include several molecular mechanisms activated during OMVs and cell interaction. Found DTX4 overexpression in cell lines treated with NTBF OMVs can be as a result of PSA mediated immune suppression or to be a consequence of the unknown bacterial proteins influence to the target cells. Stannin (SNN), a highly conserved 88 amino acid protein, is associated with cell membranes and partially localized in the mitochondria. SNN simultaneously regulates components of the MAPK and ERK signaling pathways, which are important for cell proliferation [43]. SNN was upregulated in cell lines treated with NTBF OMVs. STRAP also activates the MAPK/ERK signaling pathway and reduces the inhibitory activity of the Cdk p21. Its activity leads to phosphorylation of the retinoblastoma protein (pRb), and promotes activation of tumor cells growth and proliferation. STRAP has both oncogenic and suppressive potential, depending on the tissue type. Its suppressive function has been demonstrated in the colon [44]. STRAP was downregulated in cell lines treated with NTBF OMVs. [44]. Thus, it can be assumed that overexpressed SNN and SERPINB5 may increase cell growth and proliferation, which generally can be supported by suppressive potential of downregulated STRAP. In this combination all mentioned proteins may promote cell survival under OMVs attack [45].
The most interesting variants of proteins which were found up- and down-regulated in SW-837 cell lines treated with ETBF OMVs were: BRD7, TPM3 and prohibitin. TPM3 is involved in activation of epithelial-mesenchymal transition, with subsequent E-cadherin degradation and the following cell migration [46]. Interestingly, ETBF OMVs containing fragilysin promote the enzymatic degradation of E-cadherin. Since E-cadherin is an indirect fragilysin’s substrate, then TPM3 overexpression can be defined as a concomitant regulatory factor for modifying intercellular adhesion. The BRD7 protein, which was downregulated in the SW837 cell line treated with ETBF OMVs, is a regulator of the Wnt-signaling pathway [47]. Its direct interaction with cadherins is necessary for its inhibitory effect on tumor growth. When the BRD7 protein is downregulated, tumor growth is expected to increase. Prohibitin, a global regulator of cell proliferation and apoptosis, was downregulated in cell cultures treated with ETBF OMVs. Overexpression of prohibitin has been described as one of the possible mechanisms of transformation from adenoma to colorectal cancer. Prohibin is involved in Wnt-pathway regulation and contributes to the negative regulation of apoptosis [48]. Thus, cell lines treated with ETBF and NTBF OMVs are able to enhance cell proliferation contributing cell’s survival under bacterial vesicles attack.
A significant increase in previously observed and newly detected proteomic effects was observed in the cell lines treated with ETBF or NTBF OMVs for 5 hours. Only one of the pathways (HSA-446353 or hsa04141) was activated in each cell line treated with ETBF or NTBF OMVs for 3 hours. However, the global proteome changes had occurred in the cell lines treated with ETBF or NTBF OMVs for 5 hours. More than 30 different pathways were activated in cell culture treated with NTBF OMVs. Interestingly, the most significant signaling pathways contained the following upregulated proteins: YWHAB; YWHAQ; YWHAZ. Those proteins are the basis of cell cycle signaling pathway, Infection Disease and Immunity signaling pathway, Program cell death signaling pathway, SARS-CoV-1/ SARS-CoV-2 Infection signaling pathway and Transcriptional Regulation by TP53 signaling pathway. This group of proteins is characterized by involvement in a wide range of protein interactions, mainly affecting cell proliferation and cell apoptosis. It is known that inhibition of these proteins leads to cell apoptosis. For example, YWHAB protein plays important roles in cancer signaling. Collectively, YWHAB, YWHAQ and YWHAZ are considered to be tumor suppressors, whose down-regulation has been frequently detected in many types of cancer [49, 50]. Since upregulation of YWHAB was observed in culture treated with NTBF OMVs, it can be assumed that cells continue to receive an additional proliferate stimulus when contacting with bacterial vesicles. The involvement of proteins which are the basis of SARS-CoV-1/ SARS-CoV-2 Infection signaling pathway in the process of cell-bacterial OMVs interaction is probably a consequence of the presence of common targets/receptors for viruses and bacterial vesicles on/in the epithelial cells. In the particle, SARSCoV-1 N binds to the host protein YWHAB, which regulates nucleocytoplasmic trafficking [51].
Several signaling pathways which were characterized by involvement of mentioned proteins (YWHAB; YWHAQ; YWHAZ) were also activated in cell lines treated with ETBF OMV. Among the activated pathways were the follow: Apoptosis (HSA-109581), Immune System (HSA-168256), Infectious disease (HSA-5663205), Negative regulation of NOTCH4 signaling (HSA-9604323), Signaling by Rho GTPases (HSA-194315) and Innate Immune system (HSA-168249). Immune System (HSA-168256) and Innate Immune system (HSA-168249) pathways activated by ETBF OMVs deserve more attention because overexpressed Serpin B3, which is involved in regulation of mentioned pathways, modulates the host immune response against tumor cells and provides cells apoptosis. It has been demonstrated that dysregulation of SerpinB3, COX-2 and β-Catenin are associated with more advanced tumor stages in colorectal cancer. The in vitro results supported a driving role of SerpinB3 in the upregulation of COX-2/ β-Catenin positive loop, associated with a more aggressive cellular phenotype [52]. Previously it has been shown that Bacteroides fragilis toxin coordinates a pro-carcinogenic inflammatory pathway via targeting of colonic epithelial cells [53]. SerpinB3 probably plays one of the key roles of the formation of the complex tumorigenic reactions in the cells treated with ETBF OMVs. Fragilysin associated with ETBF OMVs can possibly contribute to the overexpression of SerpinB3 and furthermore promotes the activation of MAPK6/MAPK4 signaling pathway (HSA-5687128) [54].
IL18 is a pro-inflammatory cytokine primarily involved in epithelial barrier repair, polarized T helper 1 (Th1) cell and natural killer (NK) cell immune responses [55]. IL18 was found to be overexpressed in cell lines treated with NTBF OMVs. The secretion of a pro-inflammatory cytokine is particularly surprising as the expected effect of NTBF OMVs is to reduce the inflammatory response. However, it is known that B. fragilis capsular polysaccharide A (PSA) is associated with OMVs and delivered to intestinal dendritic cells (DCs) to induce interleukin-10 (IL-10) production. IL10 suppresses inflammation [42]. Possibly, IL-18 may play a profound role during the initiation phase of an immune response by recruiting plasmacytoid DCs and modulating the function of DC2s [56]. The recruited DCs further interact with the PSA-containing OMVs. A possible mechanism of the suppression of epithelial cell inflammation by the secretion of IL18 is shown in Fig. 7.
Increased amounts of the FLNA and HSPB1 were observed in the culture treated with EBTF OMVs. Both proteins are involved in the pathway with the following interleukin-18. But in cell lines treated with ETBF OMVs the upregulation of HSPB1 did not lead to the secretion of interleukin-18 as observed in the culture treated with NTBF OMVs. However, it can be assumed that similar mechanisms may also be active in cell culture when treated with ETBF OMVs. The main problem with a timely anti-inflammatory response is probably related to the presence of a toxin that can delay such effects by affecting the cell cytoskeleton. We hypothesized that the cell treated with the toxin is rebuilding the cytoskeleton structure and is therefore unable to respond to the OMVs attack in a timely manner. This hypothesis and the mechanism of IL18 mediated suppression of inflammation require further experimental evaluation.
{kind=link}