Ambient temperature is an important environmental cue that fluctuates momentaneous, diurnally and seasonally. Temperature is physical entity that can be seen as a gradient that ranges from freezing to heat in the so called ‘physiological’ range [1, 2, 3, 4]. The perceived temperature provides plants with valuable information to tune growth, development and phenology to its current (temperature) environment. Although extreme, i.e. stressful, temperatures can cause damage to plants and generally leads to growth cessation, or even death, acclimation of plant architecture, physiology, phenology, growth and development to mild sub-optimal cold and warm temperatures can help plants to maintain optimal performance under unfavourable (sub-optimal) temperature conditions [5, 6, 7].
Current climate change leads to more extreme and irregular weather events and increased levels of stressful temperatures, exposing ecosystems and agriculture to episodes of severe drought, flooding and heat [8, 9, 10, 11]. Independent methods consistently project climate change to have a negative impact on crop yield and already a 1°C increase in average global temperature is projected to lead to major yield losses in staple crops such as wheat, maize and rice [12, 13, 14, 15, 16]. These adverse effects coincide with an increase in global food and feed demand and poses a considerable challenge for the agricultural sector to increase food security [13, 17]. Thus, there is a pressing need for the development of thermotolerant crops that can withstand adverse temperatures.
Thermomorphogenesis; acclimation to mild warm temperatures
Acclimation to high ambient temperatures, as for instance seen in the model plant Arabidopsis thaliana when grown at 27–28°C compared to standard laboratory conditions (20–22°C), is called thermomorphogenesis [2, 18, 19, 20]. In Arabidopsis seedlings, thermomorphogenesis is characterized by elongated hypocotyls and hyponasty (upward movement) of the cotyledons [18, 21, 22]. In adult vegetative plants, high ambient temperatures result among other traits in longer petioles, hyponasty and alterations in leaf blade shape and size [5, 22, 23]. Together, thermomorphogenesis triggers an open rosette structure that is proposed to aid the plant’s cooling capacity [24, 25]. High ambient temperature also leads to early flowering (reviewed in 26). In addition to Arabidopsis, thermomorphogenesis occurs in many crop species such as tomato, wheat and cabbages [16, 19].
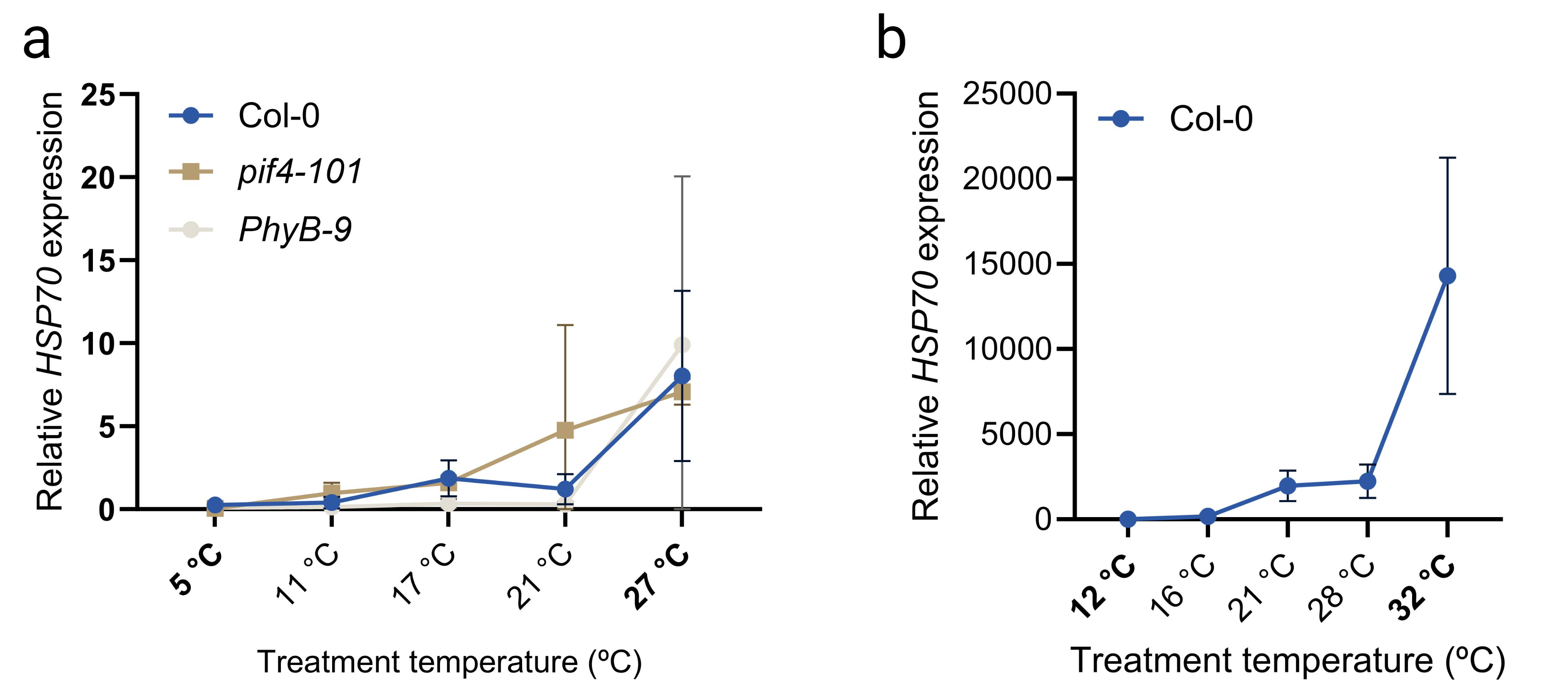
The transcription factor PHYTOCHROME INTERACTING FACTOR 4 (PIF4) is considered a central signalling hub for thermomorphogenesis [18, 27]. In response to high temperature, PIF4 levels rise and the protein binds to the promoters of auxin biosynthesis genes such as YUCCA8, thereby enhancing auxin biosynthesis at high temperatures, leading to thermomorphogenesis [18, 28]. Accordingly, pif4 null mutants lack the ability of e.g. hypocotyl elongation in response to a higher ambient temperature [29]. Another molecular maker for high temperature is HEAT SHOCK PROTEIN 70 (HSP70) as its transcription directly scales with the perceived temperature [30].
In Arabidopsis thaliana so far three bona fide thermosensory events have been described involving phytochrome B (phyB) [31], EARLY FLOWERING 3 (ELF3) [32] and PHYTOCHROME INTERACTING FACTOR 7 (PIF7) [33]. Likely, more sensory events remain to be discovered. Warm temperature-dependent conversion of the active phyB Pfr to the inactive Pr conformation leads to nuclear exclusion of phyB and release of PIF4 inhibition [28, 31, 34]. EARLY FLOWERING 3 (ELF3) is a component of the evening complex (EC) of the circadian clock that restricts PIF4 expression during the early evening [35]. Additionally, ELF3 can interact with PIF4 protein in an EC-independent manner, preventing PIF4 to activate its targets (Nieto et al., 2015). Later, it was shown that the prion-like domain of the ELF3 protein provides thermosensory input [32]. The prion-like domain causes ELF3 to form reversible aggregates by liquid-liquid phase separation at high ambient temperatures [32]. As a result, ELF3 can no longer be integrated into the evening complex, nor can function as a negative regulator of PIF4. Interestingly, temperature sensitivity of ELF3 scales with the length of the polyQ tract in the domain [32]. In the same year, it was shown that (PHYTOCHROME INTERACTING FACTOR 7) PIF7 plays an important role in regulating growth during daytime. It was found that translation of PIF7 mRNA, and thereby protein levels, are enhanced by high temperature-dependent relaxation of a PIF7 mRNA hairpin structure [33]. Alike PIF4, PIF7 can bind to promoters of auxin biosynthesis and signalling genes and likely PIF7 and PIF4 are dependent on each other during the induction of thermomorphogenesis by forming heterodimers [36]. Downstream of these signalling events, diverse chromatin remodelling, hormone-mediated signalling events and transcriptional processes have a role in translating the temperature information into proper responses to the prevailing temperature (reviewed in [2, 19, 20].
Cold acclimation
Acclimation to low, but non-freezing temperatures, is called cold acclimation. In Arabidopsis thaliana, cold acclimation occurs at temperatures between 0 and 5°C and leads to major transcriptional changes to induce physiological and biochemical modifications, resulting in enhanced freezing tolerance [37, 38]. Low temperatures amongst others triggers, growth inhibition, changes in cell wall composition to maintain cell wall integrity, capture of Reactive Oxygen Species (ROS), production of cryoprotective proteins, increased osmolyte levels and adjustments in photosynthesis [39, 40, 41, 42, 43]. Key to cold acclimation is the ICE1-CBF-COR regulon [37, 38, 44]. In response to cold the basic-helix-loop-helix type transcription factor INDUCER OF CBF EXPRESSION 1 (ICE1) binds to the promoter of C-REPEAT BINDING FACTOR (CBF) genes. In turn, the CBF genes (CBF1, CBF2 and CBF3) bind to the cis-element of the COLD RESPONSIVE (COR) genes, thereby activating their expression. This leads to the induction of cold acclimation responses, including induction of cryoprotective proteins that protect plant cells against cold stress-induced damage to the membrane [37]. ICE1 is mostly regulated at the post-translational level by protein kinases, such as MITOGEN ACTIVATED PROTEIN KINASES (MAPKs), that can either stabilize or de-stabilize the ICE1 protein by protein phosphorylation (reviewed in [4]).
Studying temperature acclimation across the temperature spectrum
The molecular regulation of thermomorphogenesis and cold acclimation is currently well understood. To the best of our knowledge however, none of the identified molecular factors have an apparent role in acclimation to both cold and warmth, despite being part of the same temperature continuum. Investigating and identifying molecular factors that regulate acclimation processes along the temperature spectrum are considered prime targets for the development of climate tolerant crops [7]. It thus is essential to study plant acclimation responses across a gradient of applied temperatures (i.e. perform temperature dose-response assays). This is not only true for responses to abiotic stresses, but also for biotic stresses, as both cold and warm ambient temperatures also have an effect on plant immune responsiveness [45, 46].
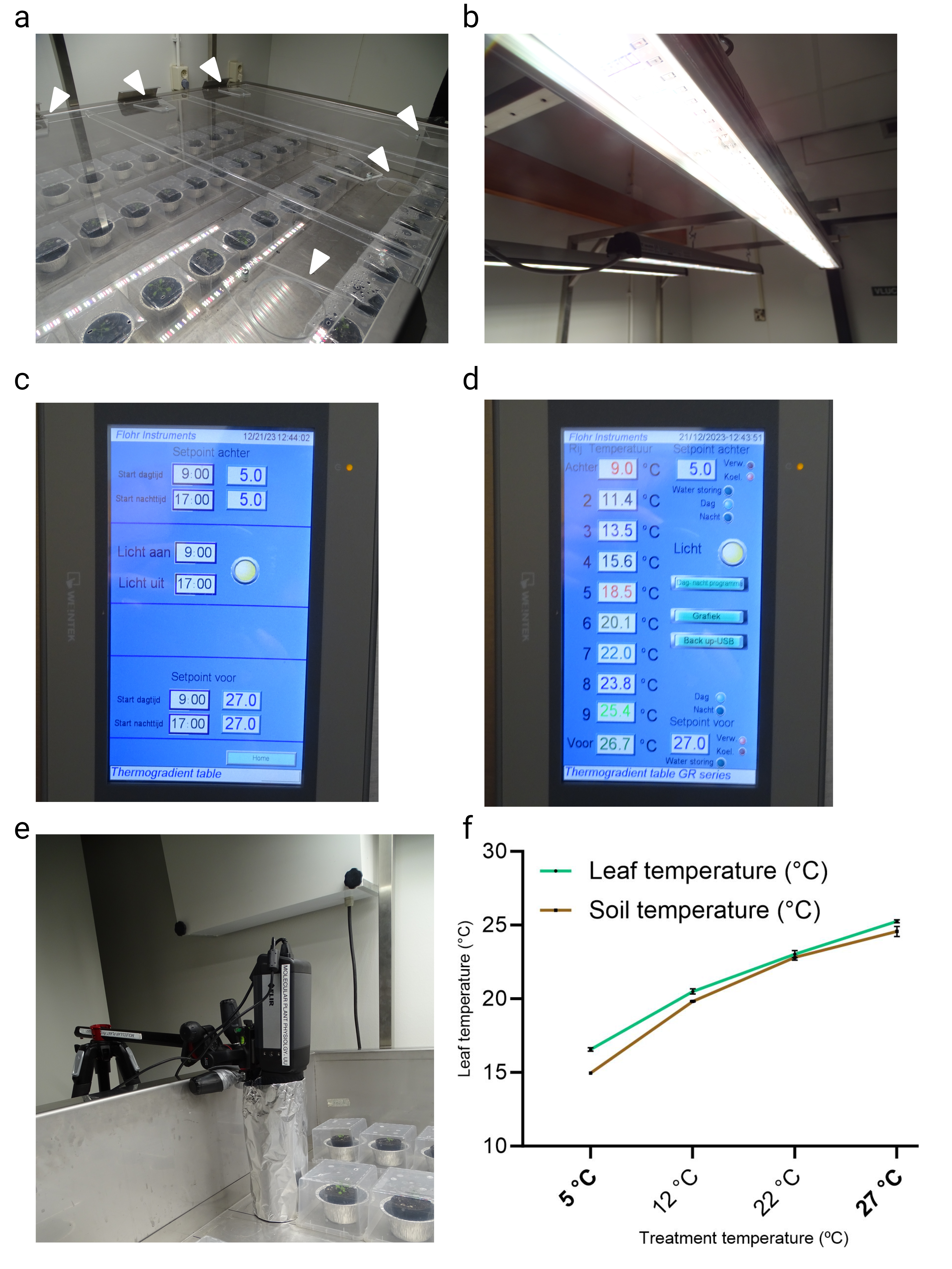
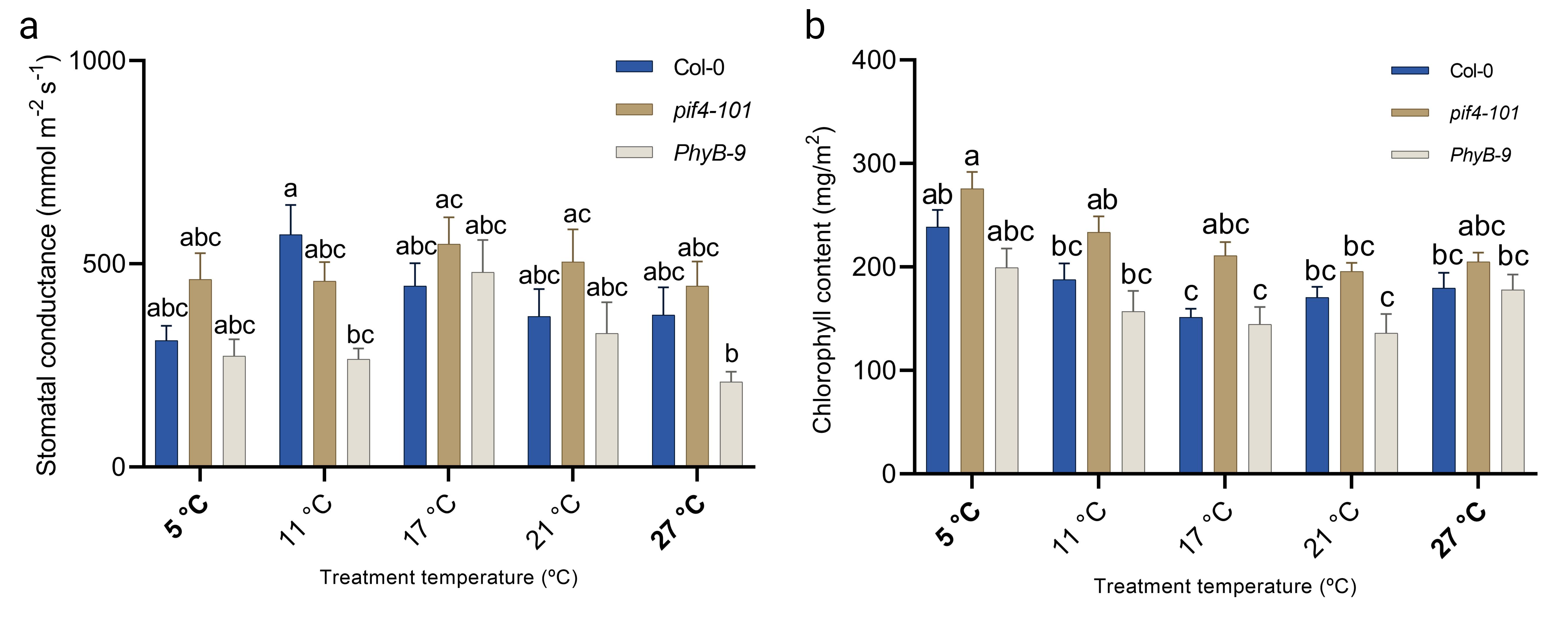
Studies that cover abiotic signal gradients are very common in ecological research [47, 48, 49, 50, 51]. Although studying spatial and temporal gradients that occur naturally can explain variation on a large scale, such studies also typically include many confounding factors such as fluctuations in precipitation, soil type and differences in, for example, wind and light exposure. Therefore, if one’s aim is to understand the change in a single environmental factor, geographical gradients, such as elevation and latitudinal gradients, are often relatively poor proxies [52]. This might be especially true when one wants to associate mild changes in temperature to growth and development, as the natural stochasticity in the temperature signal (momentaneous) and rhythmicity (diurnal and seasonal changes) both may hamper detection of causal relations between mild/small non-stochastic changes, such as consistent average climate warming, in temperature values and plant trait effect sizes. This is especially true in non- or semi-controlled field environments, where light quality and quantity and other parameters can strongly fluctuate. In the context of temperature studies, consistency in light intensity and quality is crucial, as major parts of the temperature and light signalling networks overlap and have the potential to evoke very similar responses such as, e.g. elongation of the hypocotyls and petioles [53, 54, 55, 56, 57]. Given these confounding effects, studying effects of temperature dose on plant traits thus ideally requires a stable research environment (laboratory climate-controlled rooms, cabinets or green-houses) where temperature is the only parameter that is empirically tweaked. Studying plant temperature dose-responses over a wide temperature gradient is however a technical challenge; i.e. each temperature setting would require one cabinet or growth room. To overcome these limitations we considered whether thermal gradient tables, that are commercially available and often used for e.g. assessing seed quality by breeders [58, 59], are suitable for studying plant traits over a temperature gradient within the same confined experimental set-up under otherwise controlled environmental conditions (e.g. fixed photoperiod, humidity, light quantity and quality). We describe thermodynamic and technical aspects of our thermal gradient table setup and validate the table by demonstrating that diverse typical morphological, (molecular) physiological and developmental aspects of cold acclimation and thermomorphogenesis can be recapitulated by using our set-up; including temperature dose-response effects on seed germination, hypocotyl elongation, leaf development, hyponasty, temperature marker gene expression, ion leakage, hydrogen peroxide levels, stomatal conductance, chlorophyll content and rosette growth. We provide detailed technical information, growth protocols and considerations to aid the research community in rapid incorporation of thermal gradient table systems into the research field of temperature signalling and response.
{kind=link}
{kind=link}
{kind=link}
{kind=link}