In the present study, we provided hitherto undocumented evidence suggesting that VPL-M1 projections play an important role in ameliorating CPSP in rats. We investigated the effects of optogenetic techniques to modulate the excitability of glutamate neurons in VPL-M1 projections in CPSP rats. Our findings showed that inhibiting the excitability of these neurons increased the pain threshold and ameliorated the symptoms of pathologic pain. Additionally, we assessed the effects of this inhibition on pain-related proteins (c-Fos, VGLUT2, and NR2B) by qRT-PCR and immunofluorescence. This analysis suggests that raising the pain threshold in CPSP rats may be associated with the down-regulation of these proteins in the M1 region. These findings further our understanding of the mechanisms of central sensitization in CPSP and highlight the potential of VPL-M1 projections as a target for alleviating this condition.
A highly accepted animal model for replicating CPSP in humans is the CPSP model of thalamic hemorrhage caused by injection of type IV collagenase in the VPL(De Vloo et al. 2017; Hornung et al. 2020). This model is chosen as it more closely resembles hemorrhagic stroke, a known cause of CPSP in humans(De Vloo et al. 2017). Additionally, a unilateral hemorrhagic lesion in the VPL better mimics the pain hypersensitivity experienced by CPSP patients. The thalamus, especially the VPL nucleus, is widely thought to play a key role in CPSP development(Li et al. 2021). Central pain is often observed after thalamic injury, and lesions confined to the VPL have also been shown to suffice for impairing temperature sensation and triggering CPSP in humans(Volz et al. 2012). Furthermore, studies suggest that thalamic lesions trigger CPSP only when they involve the VPL nucleus and that the ratio of CPSP to non-CPSP in humans with a VPL lesion compared to lesions of other thalamic regions is approximately 80 percent. Additionally, there is evidence that CPSP-induced nociceptive hypersensitivity is causally related to ongoing neuronal activity in the lateral thalamus(Gross et al. 2007; De Vloo et al. 2017; Zhou et al. 2018). Therefore, the rat model of thalamic hemorrhage caused by injection of type IV collagenase in the VPL region is more in line with the requirements of an animal CPSP model. We repeatedly tested various aspects of the model, including the dose, concentration, and speed of type IV collagenase injection, as well as the precise coordinates of the VPL region in the rats, to ensure its robustness and reproducibility. Finally, we validated the successful establishment of the model using rat behavioral methods.
Glutamate, the most widely distributed excitatory neurotransmitter in mammalian brains, plays a crucial role in numerous physiological functions(Xue et al. 2008). However, increased glutamate or its stimulation is thought to be a significant contributor to the pathophysiology of neurological disorders like stroke and painRecent Advances in the Modulation of Pain by the Metabotropic Glutamate Receptors(Mazzitelli et al. 2022). In CPSP patients with nerve injury, overactive neurons, CNS neuron sensitization, and an imbalance between excitatory and inhibitory functions within the nervous system lead to central sensitization(Limongi et al. 2021). Glutamate and other excitatory neurotransmitters play a major role in this process(Zheng et al. 2021). Studies have shown markedly active glutamatergic pathways in centrally sensitized nociceptive regions, accompanied by decreased nociceptive thresholds (anomalous nociception), increased nociceptive response (nociceptive sensitization), and pain spreading to non-injured areas. Previous studies suggest that failure of cortical inhibitory control leads to dysfunction of pain neural circuits associated with cortical layers, resulting in increased excitatory transmission and overactivity of VPL glutamatergic neurons(Fukuyama et al. 2018). This dysfunction is suggested to be associated with cortical deficits in pain. Our findings align with this suggestion, as optogenetic inhibition of VPL-M1 glutamatergic neuron excitability alleviates CPSP neuropathic pain symptomatology compared to the CPSP group. Considering the time-dependent nature of neuromodulation in functional recovery, we assessed the long-term effects of modulating VPL-M1 projections on CPSP. We compared pain-related behavioral changes in CPSP rats during 12 days of consecutive optogenetic intervention, recording changes in pain thresholds during and after the interventions, and analyzing effects on the expression of M1 pain-related factors c-Fos, VGLUT2, and NR2B. Additionally, to understand the VPL-M1 pain mechanism more comprehensively, we investigated the immediate effects of modulating the VPL-M1 pathway on CPSP. We recorded real-time changes in pain behavior and state when photostimulation was on and off, analyzing the direct effect on pain thresholds for direct evidence of optogenetic pain symptom improvement.
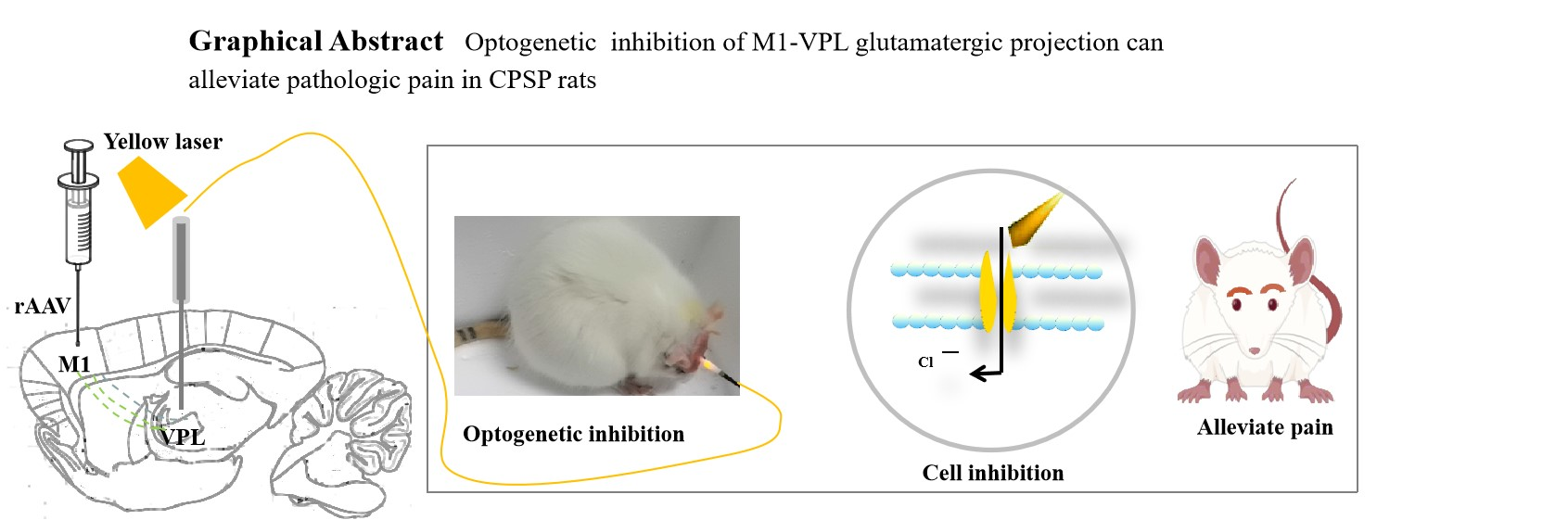
Currently, non-pharmacological treatments like neuromodulation, commonly used in CPSP, stimulate all cells in a specific area. This broad targeting can lead to side effects(Ostergard et al. 2014; Cochran et al. 2015). Optogenetics, conversely, combines light and genetic techniques to activate or inhibit specific cells, offering temporal and spatial specificity(Berglund and Gross 2020). This eliminates the side effects associated with broader neuronal targeting and holds promise as a future pain therapy(Nagel et al. 2002; Fortin et al. 2008). However, few studies have investigated its application in CPSP. In this study, we employed an optogenetic strategy to inhibit glutamatergic neurons in the M1-VPL projections to explore their potential role in CPSP pathogenesis. We first precisely injected rAAV virus into M1. After 28 days, glutamatergic neurons in M1 expressed the virus, which acted as a retrograde tracer. These transfected neurons project anterogradely to VPL. We then investigated the therapeutic effects of VPL-M1 projections on CPSP rats by laser stimulating the VPL. For expression, we chose to inject recombinant adeno-associated virus (rAAV-CaMKIIa-eNpHR3.0-mCherry-WPRE-pA) carrying the CaMKIIa promoter and encoding an inhibitory class of light-sensitive channel proteins (NpHR) into M1. CaMKIIa specifically targets excitatory glutamatergic neurons, allowing optogenetic modulation of their activity with spatial and temporal precision(Zybura et al. 2020). NpHR, the most widely used inhibitory photosensitive protein in neuroscience, activates the chloride pump and induces cellular hyperpolarization upon yellow light (589 nm) irradiation, thereby inhibiting glutamatergic neuron excitability and directly modulating pain behavior in rats(Chuong et al. 2014). Our results showed that yellow light irradiation of VPL increased both mechanical and thermal pain thresholds, improving CPSP symptoms in rats. Previous studies have demonstrated glutamatergic neuron activation after stroke, leading to excess glutamate release, increased neuronal sensitivity to glutamate, elevated intracellular ions, enhanced oxidative stress, and neuronal apoptosis(Metcalfe et al. 2022). Therefore, inhibiting overexcited glutamatergic afferent neurons may alleviate pain symptoms in CPSP rats. These findings suggest that interventions controlling the activity and output of glutamatergic neurons projected by M1-VPL could be an ideal strategy for CPSP pain management.
It is now understood that glutamate plays a crucial role in central sensitization, and targeting its system offers promising treatment options for neuropathic pain with better efficacy and fewer side effects(Grigoras and Stagg 2021). VGLUT2, responsible for glutamate transport and release, is increasingly linked to neuropathic pain. Its abundance in key pain pathways like the spinal cord, thalamus, and brainstem suggests its involvement. As expected, VGLUT2 knockout in mice significantly reduces pain symptoms, aligning with our findings(Wang et al. 2019). NR2B, a key subunit of the glutamate receptor NMDA, plays a vital role in pain regulation. Studies have confirmed its higher expression in forebrain regions like the cortex(Wang et al. 2019). Since glutamate is excitotoxic, its elevated levels after central nervous system damage bind to receptors, triggering excitotoxic effects. We observed increased NR2B expression in the M1 region of CPSP rats compared to normal rats. Down-regulating NR2B expression in this region also suppressed pain hypersensitivity and increased pain thresholds. The c-Fos protein, a marker of neuronal activity changes, is widely used to study analgesic mechanisms(Wu and Zhuo 2009). Studies have shown increased c-Fos in pain-induced rats with hypersensitivity, relieved by c-Fos expression inhibitors(Santos et al. 2018). Similarly, our study showed c-Fos overexpression in the M1 region of CPSP rats. Optogenetic inhibition of M1-VPL glutamatergic neurons down-regulated c-Fos expression and alleviated pain. We hypothesize that this mechanism involves hyperpolarization of glutamatergic neurons and blocking voltage-dependent calcium channels, leading to reduced calcium influx and preventing c-Fos expression induction, ultimately producing analgesia(Zhou et al. 2022).
Our study has several limitations. First, the invasive optogenetic technique used could not avoid nerve damage during surgery. Therefore, there is an urgent need to improve photogenetic technology for clinical application, addressing the challenges faced in translation. Second, CPSP patients not only experience thalamic hemorrhages but also hemorrhages or cerebral infarcts in other brain regions, with both ischemic and hemorrhagic causes. Additionally, varying lesion sizes and locations contribute to differences in underlying pathophysiology. Therefore, further investigation of more ideal CPSP animal models is necessary. Third, using only adult male rats in our in vivo model potentially introduces gender bias, as studies suggest differences in pain response between genders. Additionally, the type IV collagenase-induced VPL hemorrhage model, while useful, is distinct from the human CPSP model and has limitations in behavioral testing. These factors may affect our analysis of pain behavior.
{kind=link}