Disentangling the role of Notch signalling during early embryogenesis is critical to understanding the mechanisms involved in tissue formation and boundary establishment and refinement. The processes involved in tissue and boundary formation are conserved in later stages of the organism’s life as in stem cell renewal and differentiation. Unraveling the interactions between different signalling pathways such as Notch signalling and Toll signalling is also important for understanding disease pathogenesis, where the same processes are misregulated and lead to disease26.
The role of Notch signalling during embryogenesis has long been appreciated in the context of neurogenesis and myogenesis, which both occur after early mesoderm development27,28. However, while there’s evidence to suggest that it is active in the mesoderm, its role here is largely unexplored. There have been very few genetic links characterized between Notch and genes expressed in the mesoderm, although these links are observed in adult tissues or during neurogenesis and myogenesis during later stages of embryogenesis and larval stages29,30. Notch has been shown regulate the expression of; Twi, Asph, and string29,30. Here, for the first time, we present evidence of its activity in the ventral presumptive mesoderm and identify target genes, which are critical to mesoderm formation and gastrulation, that are dependent on Notch signalling for their expression. Among these genes are; tinman, Mes2 and Mef2 which all encode transcription factors that are necessary for myogenesis during embryonic and larval stages31,32,33,34,35,36. Some of the genes, whose expression was dependent at least in part on Notch signalling were factors that are involved in orchestrating or facilitating cell shape changes that are necessary for mesoderm invagination and internalization37. These include; Traf4, which encodes a cytoplasmic protein involved in TNF signalling, which plays a critical role in the cellular apical constriction, which is necessary for ventral furrow invagination37. Another affected gene, Neurotactin, encodes a cell adhesion molecule38. Decreases and increases in Notch signalling also affect the expression of htl and stumps, which encode key players of FGF signalling, which is required for mesoderm spreading8. Here we show that Notch signalling is required for the expression of Stumps but negatively regulates htl expression. Although the loss-of-function mutants did not show defects in mesoderm spreading when cross-sections were examined. This indicates that expression of these genes is only partially dependent on Notch signalling, which is consistent with previous studies that characterized them as target of Twist34.39,40. Since Twi protein is expressed in these loss-of-function Delta mutants, this indicates that the Twi protein reductions observed are not detrimental and are sufficient for some downstream target gene expression and therefore do not perturb mesoderm spreading.
To test the sufficiency of Notch signalling to drive the expression of mesodermal genes, we developed a novel light-gated tool that allows precise temporal control. This system provides several advantages compared to traditional overexpression models that have been used previously; 1) The system is titratable as shown by the incremental expansion of sim expression (Fig. 3), 2) the combination of this system with tissue-specific gal4 drivers can provide tissue specific and temporal control, and 3) it has the potential to allow precise spatial control if combined with light sheet microscopy. For our experiments, it allowed us to have temporal control of activation of the system according and to titrate Notch activity. There are a few cells in the embryo that did not express sim in regardless of photoactivation time, which indicates that there are additional regulatory mechanisms that either directly repress sim or indirectly by inhibiting NICD translocation to the nucleus.
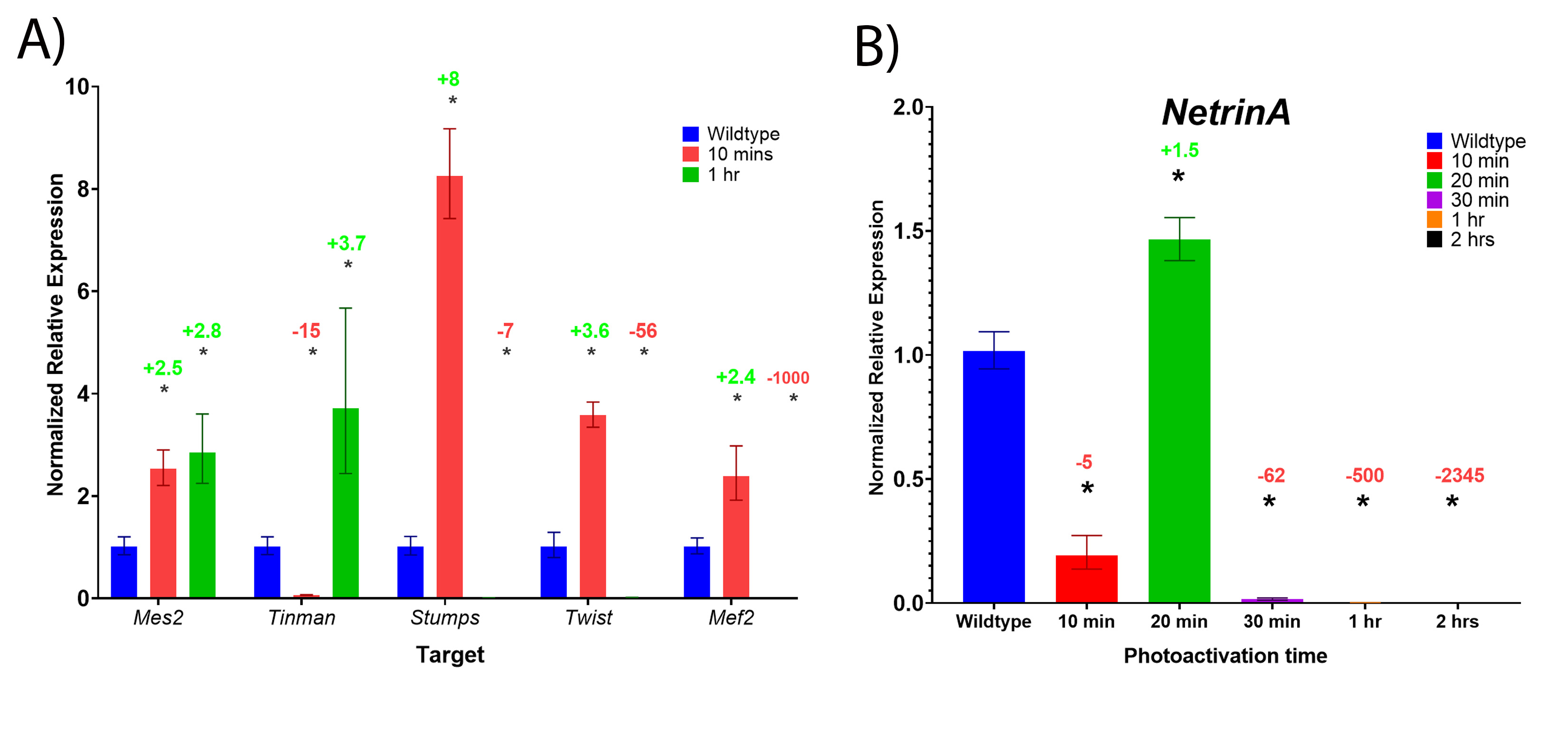
In this study, we present evidence that critical mesodermal genes require Notch signalling for normal expression, shown by significant decreases in mesodermal gene expression in both loss-of-function and gain-of-function Notch mutant embryos. In the gain-of-function Opto-Notch mutants that were photoactivated for a longer duration, expression of most of the mesodermal target genes was significantly reduced (Fig. 4), however expression in those photoactivated for shorter durations was significantly increased for many of the candidate genes including; Mes2, Tinman, Stumps, Twist, Mef2, and NetA (Fig S2). This indicates that Notch is sufficient to increase their expression, and that different genes require different levels of Notch signalling. The decrease in expression after a long duration of photoactivation is likely due to two reasons; 1) negative feedback inhibition inputs that follow increases in gene expression, some of which from Notch target genes encoding gene repressors, (E(spl)C and potentially emc)29 and 2) decreases in Twist expression, as most of these genes are also transcriptional targets of Twist34. Our findings are supported by previous experiments preformed later in gastrulation that showed that Notch signalling regulates Twist expression directly through activating expression and indirectly by activating the expression of repressors of Twist29.
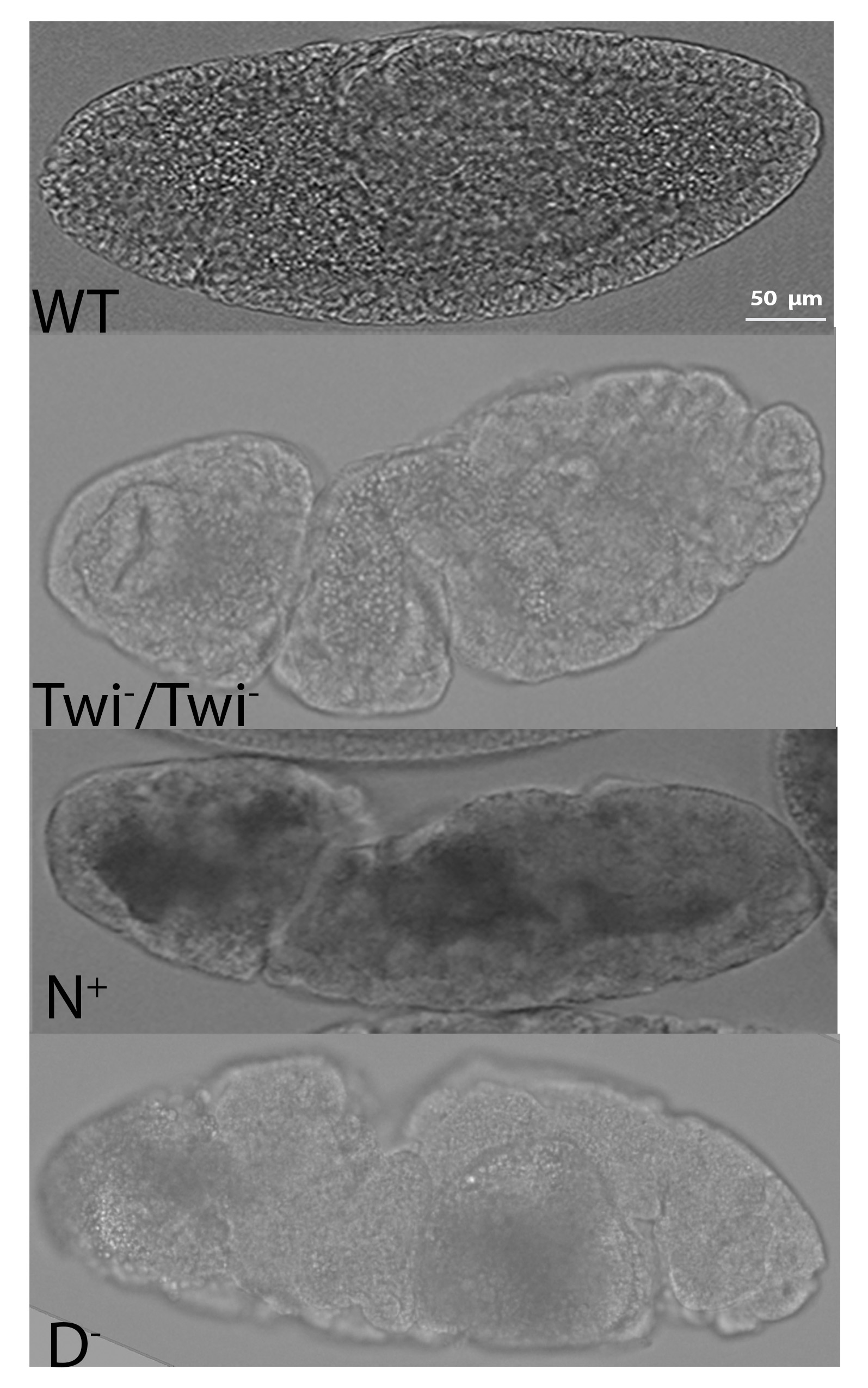
Some of the mesodermal genes were downregulated following Notch overactivation regardless of photoactivation duration including; Asph, Neurotactin, Traf4, and Heartless. Although these genes contain consensus Su(H) binding sites in their extended gene regions, our expression analysis indicates that Notch signalling is not sufficient to increase their expression in the mesoderm. This is likely due to their repression by other Notch target genes such as the E(spl)C genes, such as Emc, although this has not been tested here. Another likely mechanism is negative feedback through post-translational modifications of the endogenous NICD and Opto-NICD proteins that lead to their degradation41,42. This is supported by the decrease in fold-change of sim expression in the embryos that are photoactivated for 2 hours compared to 1 hour (Fig. 3). Put together, these findings provide clear evidence that Notch is normally active in the mesoderm during early embryogenesis and is required for normal levels of mesodermal gene expression. This requirement of Notch signalling is emphasized by the defects observed in later stages of gastrulation (after the internalization of the mesoderm) of loss-of-function mutants (Fig. 5).
Interestingly, we identified one gene, string, that behaves as a classic direct target of Notch signalling; its expression is significantly reduced in the absence of Notch signalling and is significantly increased upon overactivation of Notch signalling. String encodes a phosphatase, cdc25/ String, that activates a mitotic kinase, cdk143,44. Zygotic cell division is dependent on stg expression42,43. Loss-of-function mutants exhibit cell cycle arrest after nuclear cycle 14, which is the last cycle controlled by maternally deposited String, however, cell shape changes and morphogenetic movements are not affected except that cell numbers in the mesoderm are reduced43,44. The precise temporal expression of string has been shown to critical for the coordination of cell division43,44. The morphological defects observed in the N+ mutants described here may also be due to premature accumulation of string, causing premature cell division prior to mesoderm internalization.
Interestingly, we identified a gene, WntD, that is differentially regulated by Notch signalling across neighbouring tissues; the mesoderm and mesectoderm. Our expression analysis of loss-of-function mutants indicates that Notch signalling is required for WntD expression in mesectoderm cells that flank mesoderm cells on both sides, while it negatively regulates its expression in the mesoderm, as seen by the increase in expression in this region in the absence of Notch signalling activity (Fig. 2). The presence of Su(H) consensus binding sites in the WntD genomic region indicates that Notch can activate its expression directly by binding to Su(H). This is likely the main input on WntD in the mesectoderm. Our results imply that Notch activates the expression of a repressor of WntD in the mesoderm and in the absence of Notch signalling, this repression is relieved. The identity of this repressor remains unclear. In the gain-of-function mutant expression of WntD is expanded laterally beyond the mesectoderm, much like that of sim in those embryos and the boundary between the mesoderm and ectoderm becomes much less defined. This finding highlights the role of Notch signalling in defining the boundary between different tissues. Increases or decreases in Notch signalling, results in changes in the expression profile of these boundary cells. In the loss-of-function mutants, they lose expression of WntD and sim, while in GOF mutants, the are no longer distinct from neighbouring cells, as they express these two genes at similar levels to the neighbouring mesoderm and ectoderm cell. These changes of expression profile most likely contribute to the defects in the embryonic midline observed in the GOF mutants.
WntD encodes a secreted ligand of the Wnt family of proteins and has been identified as an inhibitor of Dorsal25. It has been shown that its expression is regulated by the Dorsal-Twist-Snail genetic network, and provides negative feedback by competitively binding to the Toll receptor instead of Spz and preventing Dl nuclear transport45. Our finding that increases in WntD expression coincide with decreases in the size of the region with the most concentrated Dorsal protein supports previous findings45. As expected, the decrease of this region, where Dorsal is most highly concentrated results in decreases in Snail- and Twist- positive cell numbers. This indicates that the mesoderm region is narrowed in both loss- and gain-of-function mutants, which emphasizes the role of Notch signalling in the development of the mesoderm. Put together, these findings provide evidence that in addition to functioning in a cell-autonomous manner to drive mesodermal gene expression, Notch signalling also provides feedback inhibition signals on the upstream patterning programme, regulated by Toll signalling through Dorsal, indirectly by regulating WntD expression.
{kind=link}
{kind=link}