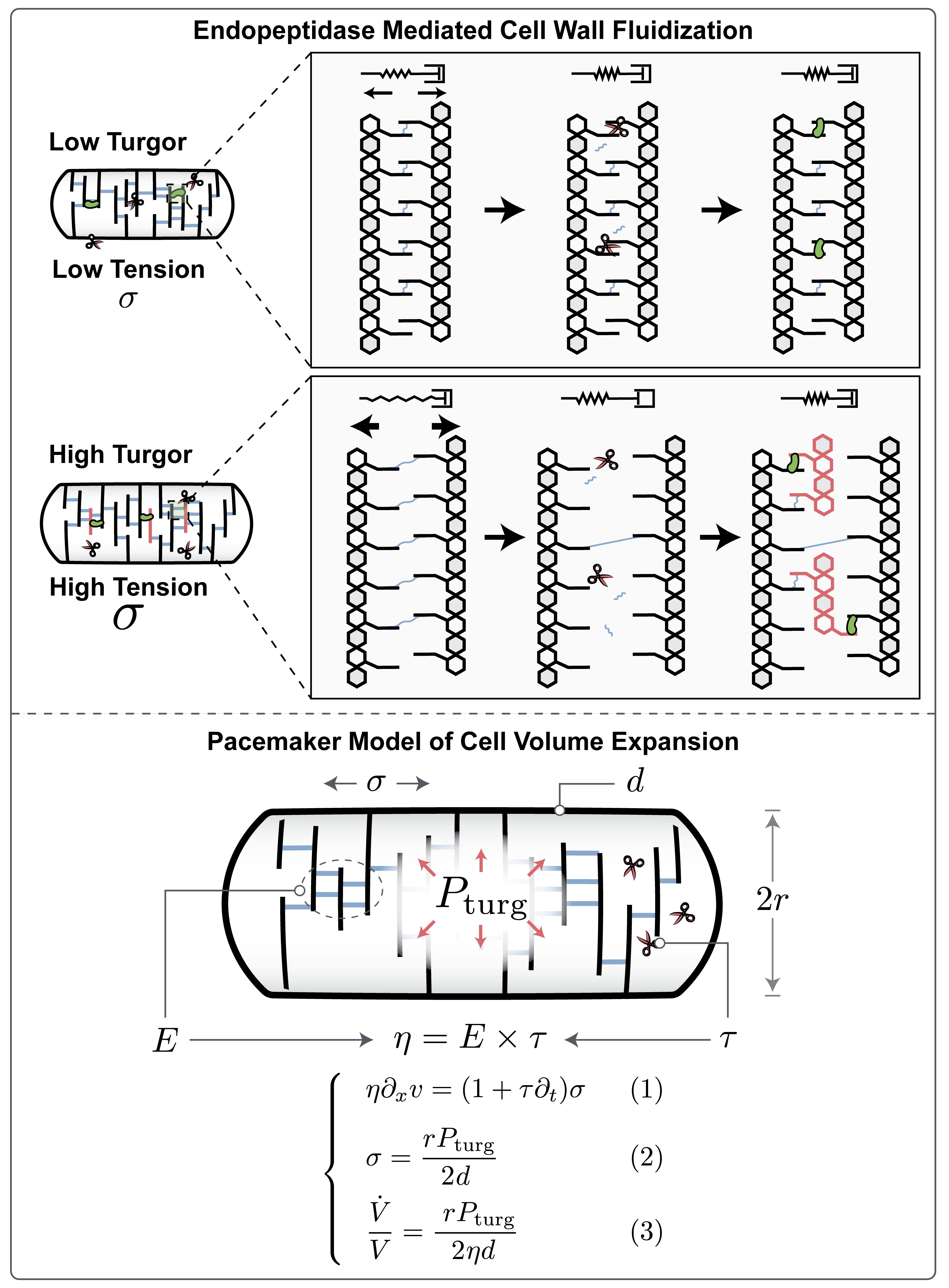
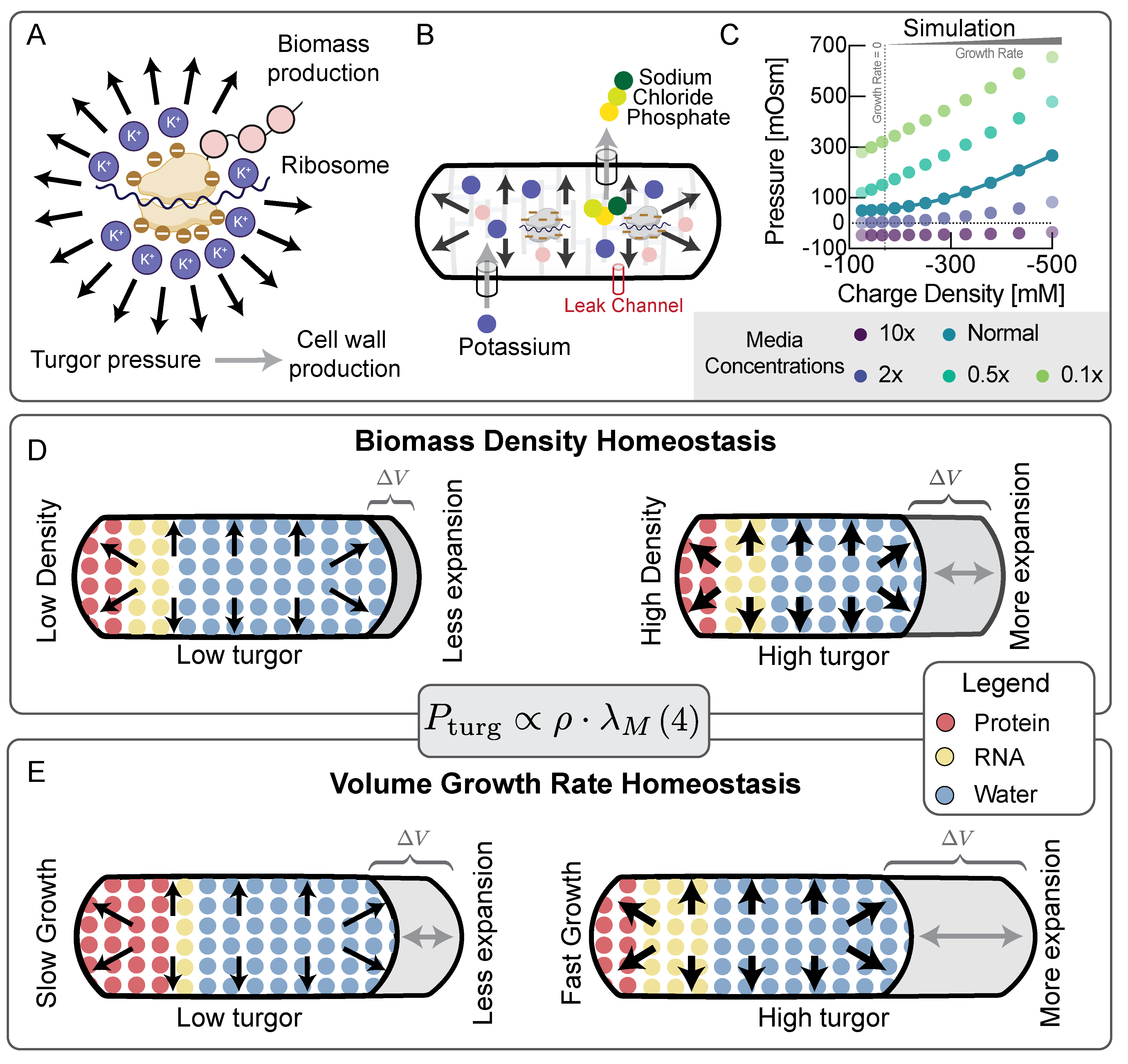
So far, our results indicate that an increase in turgor pressure with growth rate (Fig. 1) is indeed crucial for homeostasis of biomass density across growth rates (Fig. 2), enabled by endopeptidase-mediated cell wall fluidization (Fig. 3). But how is this increase in turgor pressure controlled and regulated? Turgor pressure is an osmotic pressure, directly determined by molarities of osmolytes (Fig. 5A). Potassium is by far the most abundant cellular osmolyte in E. coli with estimated concentrations surpassing the combined concentrations of all other intracellular osmolytes 13 and presumably is the biggest contributor to turgor pressure. To test if the observed increase in turgor pressure originates from changes in potassium concentrations, we measured intracellular potassium on different substrates using mass spectrometry. Indeed, we found a striking linear increase in intracellular potassium with growth rate (Fig. 5B).
These data suggest that the observed increase in turgor pressure (Fig. 1) is likely due to an increase in intracellular potassium. But how does the cell regulate intracellular potassium in precise coordination with growth rate? Potassium is positively charged, while the net charge of the cytoplasm must be neutral 27,28. Therefore, growth-rate dependent potassium concentrations must be balanced by an equivalent negative growth-rate dependent charge concentration in the cytoplasm. The concentration of the major intracellular anion glutamate has been shown to be roughly constant across growth rates 29 and thus cannot account for potassium charge balance across growth rates. However, an even bigger contributor to cytoplasmic charge comes from the potassium ions necessary to balance cellular RNA, as each nucleotide carries a negative elementary charge. Moreover, cellular RNA content as a fraction of biomass composition is well-known to increase with increasing growth rates, as measured by RNA to protein ratio 26. This is largely the result of an increase in ribosomal RNA. When we converted the RNA to protein ratios across growth rates, measured by Scott et al. 26, to an intracellular charge concentration by using our own measurements of biomass density, combined with earlier measurements of total protein and total dry mass across growth conditions 12 (see Fig. S9), we indeed found that the resulting charge concentrations presented in Fig. 5C, were sufficient to account for the observed growth-rate dependent increase in turgor (Fig. 1C). We also estimated the net charge of ribosomal proteins and confirmed that their net charge constitutes a tiny fraction of the net charge from ribosomal RNA (Table S1).
Based on these data, we propose that an increase in turgor pressure in responses to an increase in growth rate is mediated by the increase in ribosomal RNA and the corresponding retention of potassium in the cytoplasm as RNA counterions, dictated by the requirement for charge balance. We previously proposed that biomass counterions may contribute to dry mass density homeostasis 12 and we realized that the contribution from ribosomal RNA, shown in Fig. 5C, far outweighs the contribution from other biomass components in E. coli. To test this hypothesis experimentally, we asked how a change in cytoplasmic net charge is reflected in cellular biomass density. If charge balance were indeed the key determinant of turgor driving envelope expansion, then overexpressing large quantities of proteins of different net charge should affect charge balance and be directly reflected in changes in cellular biomass density. More positively charged proteins should result in a larger increase in biomass density. To test this prediction, we used an established system for expression of large quantities of useless proteins 30 to overexpress proteins of different net charge, including positively and negatively supercharged versions of GFP, developed by the Liu lab 31. Indeed, we find that biomass density continuously increases with the net positive charge per amino acid of the overexpressed protein (Fig. 5D & Fig. S10), as expected if turgor were controlled by charge balance.
{kind=link}
{kind=link}