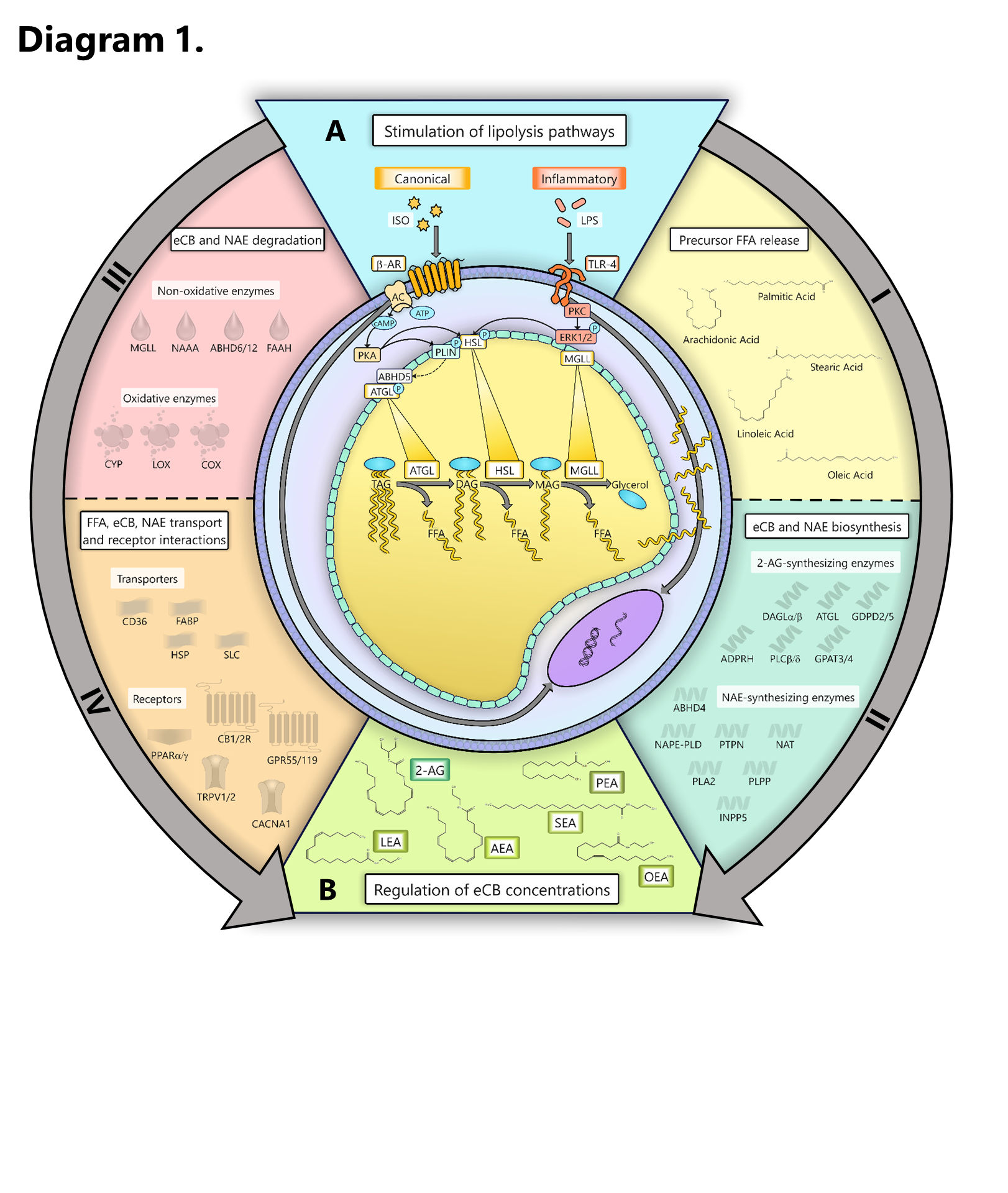
In mammals, the ECS is a potent modulator of metabolic and inflammatory processes which is widely distributed throughout organ systems, including the AT of dairy cows [17, 19, 21]. While drastic shifts in eCB and NAE concentrations and ECS component expression occur within the AT and plasma of PP dairy cows, the factors which drive these changes and their physiologic implications remain largely unknown [17, 21, 49]. Previous studies in rodents and humans have revealed that eCBs are produced within AT and modulate adipocyte FFA mobilization by way of ECS receptor signaling [50]. The present study demonstrates for the first time, however, that lipolysis pathways selectively enhance adipocytes’ production of specific eCBs, which may have significant implications for metabolic and inflammatory responses in PP dairy cows. Through lipidomic and transcriptomic analyses, we identified several enriched pathways and connections between lipolysis pathways and eCB biosynthesis, degradation, transport, and ECS signaling in dairy cows’ adipocytes.
Stimulation of lipolysis pathways enhances endocannabinoid production by adipocytes
Our data confirmed that, in bovine adipocytes, lipolysis upregulates the overall production of eCBs and NAEs however, the abundances and profiles of each are dependent upon which lipolysis pathway is activated (Fig. 1) [14, 21]. Previous studies have demonstrated strong associations between eCB and NAE concentrations and the abundance of FFA substrates in the diet and plasma, suggesting that precursor availability may play a role in their biosynthesis [51–53]. However, the role of lipolysis on eCB and NAE production had not been previously explored. Our data reveals a strong link exists between lipolysis and the biosynthesis and release of eCBs and NAEs by adipocytes.
Our results indicate that adipocytes’ production of 2-AG is augmented following the stimulation of canonical but not inflammatory lipolysis
(Fig. 1). Given the compound’s known involvement in the upregulation of feed intake, nutrient utilization, and regulation of lipolysis, enhanced levels of 2-AG may serve as a mechanism to counter negative energy balance and AT inflammation in near-calving cows, although much remains unknown in this regard [17, 54]. In contrast, our data demonstrate that the biosynthesis and release of AEA are enhanced following inflammatory–but not canonical–lipolysis. Previous studies established that immune cells biosynthesize and secrete AEA in response to LPS; however, the present study’s results are the first to evidence that a similar response is observed among adipocyte populations [55, 56]. Unlike canonical lipolysis, stimulation of the inflammatory pathway was not associated with reductions in LEA biosynthesis. These observations align with findings in human macrophages by Melis et al., who not only reported a similar increase in the release of AEA upon LPS exposure, but also proposed that, in additional to availability of precursor FFA (LA and AA), the production of AEA relies upon the degree of inflammation in the extracellular environment [57]. Taken together, these results suggest that the presence of LPS upregulates AEA production. Importantly, AEA concentrations may play a role in regulating the onset and chronic inflammation in AT [58], warranting further investigation into adipocytes’ capacity to synthesize and secrete the molecule, especially in cows with endotoxemia or septicemia.
As previous studies demonstrated that NAE abundances are reflective of cellular FFA profiles, we anticipated that induction of lipolytic pathways would enhance adipocytes’ production of NAEs. However, the results of our study indicate that adipocytes’ production and release of OEA, PEA, and SEA are reduced during canonical lipolysis. While beyond the scope of the present study, several conditions may alter the availability of FFA precursors for the biosynthesis of eCBs and NAEs in cows’ adipocytes. For example, in the AT of PP cows undergoing extensive remodeling or inflammation, adipocytes contain altered glycerolipid, glycerophospholipid, glycosphingolipid, and sterol lipid profiles whose acyl chains may serve as substrates for eCB and NAE synthesis [6, 59, 60]. In addition, lipid contents vary widely between differentiating and mature adipocytes and amongst AT depots in rodent and human models [61]. Adding to the complexity of the ECS landscape in AT, certain FFA may be preferentially incorporated into adipocytes’ membrane PL, thus, altering their availability [62]. While presently unknown, future studies should trace FFA released during lipolysis and evaluate their metabolism into eCBs and NAEs. The present findings underscore the complexity of networks which govern eCB and NAE production and release and emphasize the need to characterize the involvement of additional regulatory mechanisms, such as transcription of key genes.
Canonical and inflammatory lipolysis pathways differentially upregulate the transcription of endocannabinoid synthesizing networks in adipocytes
The transcriptional patterns observed in adipocytes provide novel insight on how canonical and inflammatory lipolysis pathways regulate eCB and NAE profiles in bovine AT (Fig. 3). Corresponding with upregulations in 2-AG release, our data indicates that canonical lipolysis upregulates adipocytes’ expression of key genes associated with the biosynthesis of 2-AG, including GDPD5 and GPAT4, which convert lysophospholipids (2-acyl lysophosphatidic acid and 2-acyl lysophosphatidylinositol) directly into 2-AG, and INPP5F, which cleaves phosphatidylinositol 4,5-bisphosphate to free phosphatidylinositol, an intermediate substrate, within the cell membrane [63, 64]. Despite enhancing the transcription of the AA-freeing enzyme PLA2G4A, LPS reduced the release of 2-AG and expression of the 2-AG biosynthesizing (and key lipase-encoding) enzyme, PNPLA2, suggesting bovine adipocytes may exhibit a reduced capacity to synthesize the metabolically active compound during endotoxemia.
The results of our present experiments indicate that, in bovine adipocytes, the production of AEA is upregulated in LPS (Fig. 3). At the transcriptional level, adipocytes’ expression of PTPN22, which cleaves phospho-AEA to AEA, is upregulated upon exposure to LPS [55]. These distinct shifts in eCB release coupled with correspondent changes in transcription suggest that, during inflammatory conditions, AA may be preferentially diverted toward AEA and away from 2-AG biosynthesis. In rodent models, AEA limits TLR-4-mediated release of inflammatory cytokines by immune cells, including TNFα, nitric oxide, IL-1β, and prostaglandins, highlighting the molecule’s capacity to attenuate inflammation [65–67]. While AEA’s anti-inflammatory effects are primarily attributed to its binding and activation of CB2R, its biosynthesis may, in addition, directly limit the amount of free AA available for conversion into pro-inflammatory mediators [11, 68]. Such effects may be particularly beneficial in cows challenged with endotoxemia, wherein AT inflammation may otherwise contribute to dysregulation of lipolysis and predispose cows to disease in subsequent lactations [69, 70].
Lipolysis pathways modify endocannabinoid-degrading enzyme expression and inflammatory markers in adipocytes
Under inflammatory conditions, eCBs may be rapidly oxidized by COX, LOX, CYP450, or hydrolyzed by MGLL in AT. Importantly, byproducts of eCB oxidation include the bioactive lipid-based mediators of inflammation known as oxylipids, which are associated with increased disease risk in dairy cows [71]. While the affinities of specific enzymes for eCB substrates remain unknown, COX have been shown to display high affinities for AA-based substances [68]. The present results indicate that, when adipocytes are exposed to LPS, their transcription of PTGS2 (COX-2) and CYP27B1 are elevated, although 2-AG levels remain comparable to basal conditions and AEA release is enhanced (Fig. 4). While the degradation of eCBs may result in the production of pro-inflammatory compounds, the direct implications of lipolysis pathway activation on eCB degradation and oxylipid biosynthesis remain undefined and should be further evaluated through silencing of degradative genes and targeted lipidomic analyses. Additionally, the results of our study indicate that adrenergic stimulation of adipocytes does not upregulate the transcription of NAE-degrading enzymes. Collectively, these findings suggest that lower NAE abundances in adipocytes’ extracellular environments may be due to reductions in the compounds’ biosynthesis or transport under these conditions rather than their oxidation or hydrolysis.
Canonical and inflammatory lipolysis regulate the expression of free fatty acid and endocannabinoid transporters
During lipolysis in PP cows’ AT, FFA are preferentially exported out of the cell, into circulating blood, and partitioned to the mammary gland for milk synthesis [6, 72]. This process is facilitated by the activity of FFA transporters, which may also bind and transport eCBs and NAEs. Known transporters of both FFA and eCBs include fatty acid translocase (CD36), fatty acid-binding proteins (FABP3 and FABP7), heat-shock proteins (Hsp-90), solute carriers (SLC27-), and sterol carrier protein-2 (SCP2) [73–75]. Binding of FFA precursors to carrier proteins may alter their availability for eCB or NAE biosynthesis, and the susceptibility of the compounds to oxidative or hydrolytic degradation. The present study revealed that, during canonical and inflammatory lipolysis, adipocytes’ transcription of FABP3 and FABP7, which are associated with the intracellular transport of lipids, are enhanced (Fig. 5). Upon adrenergic stimulation, adipocytes’ transcription of CD36 is upregulated, corresponding with heightened release of 2-AG. Coinciding with elevations in AEA abundance; however, inflammatory lipolysis upregulates the transcription of HSP1A1 and SCP2, providing further mechanistic evidence for the connection between AT inflammation and adipocytes’ release of AEA. At present, the specificities of certain FFA, eCB, and NAE transporters for various substrates are unknown, however, these proteins may play key roles in the modulation of AT ECS activation and should be further investigated in future studies.
Canonical and inflammatory lipolysis pathways differentially modulate the expression of endocannabinoid system receptors in bovine adipocytes
In the present study, adipocytes’ expression of PPARG–known as the master regulator of adipogenesis–was elevated during canonical, but not inflammatory, lipolysis (Fig. 6). This finding underscores that, within the AT of otherwise healthy cows in negative energy balance, excessive FFA mobilization may be offset through the upregulation of adipogenesis and structural remodeling of AT [6]. In contrast, the AT of transition cows challenged with bacterial infections may exhibit a reduced capacity for adipogenesis, contributing to the ectopic accumulation of FFA in AT, the blood, and the liver and progression of metabolic diseases in these animals [76]. Furthermore, our data revealed that LPS upregulates the transcription of the transient receptor potential channel, TRPV3, in adipocytes. This receptor, whose stimulation is associated with Ca2+ influx and suppression of adipogenesis, may contribute to dysregulated lipolysis and inflammation – particularly in the AT of cows afflicted with endotoxemia or septicemia [77]. These observations, largely centered around the regulation of adipogenesis, emphasize ECS receptors’ potential as therapeutic targets for PP cow health and frame the need to advance knowledge in this area.
Limitations and additional considerations
While the present study elucidated the roles of lipolysis pathways in adipocytes’ biosynthesis and release of eCBs, this model does not mimic the AT environment in vivo. Several AT components–such as nerve fibers, immune cells, and non-adipocyte-derived hormones (e.g., insulin and norepinephrine)–may influence ECS signaling, adipocyte lipolysis, and eCB concentrations in cows’ AT [17]. For example, CB1R activation on peripheral sympathetic nerves has been shown to inhibit their release of norepinephrine, which may modulate the intensity of lipolysis exhibited by adipocytes [78]. In addition, treatment of adipocytes with ISO, a pan-β-AR agonist, does not permit the isolation of effects of specific β-AR isoforms on eCB production. Therefore, to fully elucidate the involvement of β-AR in eCB synthesis and transcriptional regulation in adipocytes, future studies should consider targeted activation of β1-, β2-, and β3-AR with selective agonists. On the other hand, LPS may regulate eCB concentrations and ECS signaling within AT independent of lipolysis. For instance, exposure of AT resident immune cells (such as macrophages) to LPS is known to modulate their secretion of eCBs [79]. While beyond the scope of the present study, the complex interplay between adipocytes, other cell types, and inflammation within cows’ AT on the modulation of ECS signaling warrants further investigation.
{kind=link}