Finding resistance sources and genetic loci for marker-assisted breeding is crucial because most temperate wheat cultivars are susceptible to STB. Bread wheat accessions from India and Bangladesh have not been screened for STB, so this study identifies new sources of STB resistance. Our findings revealed complex genotypic and environmental interactions, quantitative STB resistance inheritance, and novel QTNs for STB breeding program.
Genetic variation for STB resistance
This study found polygenic control of STB severity due to its near-normal distribution over three years. For future wheat breeding and improvement, phenotypic evaluation showed significant genetic variability among tested wheat genotypes for STB resistance. The broad sense heritability estimated across environments was high, as suggested by previous studies. [18,22,23,24,25]. However, ANOVA showed significant year- and genotype-by-year effects, supporting the need for multiple-year germplasm evaluation to identify stable STB-resistant genotypes [25,26].
Many lines from the cross CHIR7/ANB//CHIR1 also showed increased resistance, highlighting the significance of the parents in this cross for the STB resistance breeding programme. STB resistance is negatively correlated with PH and DH, which are among the most confounding factors. Several studies reported increased disease severity in earlier heading and dwarf genotypes [22,27]. The relationship suggests that genotypes with tall and late phenology may escape STB with reduced infection, or that their genes may have pleiotropism or be tightly linked to STB genes. Many GWAS and QTL mapping studies have found genetic linkage as the cause of the relationship [28,29]. We found genetic factors supporting the association between STB and DH in QTNs Q.STB_DH.2A.1, Q.STB_DH.3B2, Q.STB_DH.5A.3, and Q.STB_DH.5D.4.
Association analysis and candidate genes for STB resistance
In the current study, the ‘year’ effect exerted a significant influence on disease expression, which was also observed in other studies on STB resistance, justifying the importance of multiple field trials. Of the 99 identified QTNs, only 21 were significant in more than one environment and most were of minor effects. This agrees well with the fact that resistance against STB in field experiments is quantitatively inherited trait [19,30]. Stacking favourable alleles from multiple loci, a prerequisite to developing STB resistance germplasm, is possible via introgressing the alleles from their respective donors in the present SAMP.
Of the 21 repeatable STB QTNs detected, allele A on Q.STB.2A.3, Q.STB.2B.4, Q.STB_DH.3B.2, Q.STB.6A.; C, G and T alleles on QTNs- Q.STB.5A.1, Q.STB.5B.1 and Q.STB.5B.3 were associated decrease in STB severity and displayed a significant reduction in STB severity. The allele G associated with the SNP 1122319 on chromosome 5B (Q.STB.5B.1) had the highest allele frequency (94.77%), followed by allele A associated with SNP 1059080 (88.70%) on chromosome 3B (Q.STB_DH.3B.2) and contributed significantly for reduction in the pycnidia development (Table S8). The above seven QTNs lies in the previously reported QTLs for STB resistance confounding to regions of MQTL14 (QTL identified at seedling and adult stage) on 3B and QTL10 (QTL identified at seedling stage) on 5B [19].
Some tagged QTNs concurred with the known mapping locations for STB resistance genes (Fig.7). For example, the markers of Q.STB.1D.1 was positioned at similar chromosomal region as STB resistance genes QStb.ipk-1D and close to Stb10; Q.STB_DH.3B.2 and Q.STB.3B.1 was positioned at MQTL14; Q.STB.5B.1, Q.STB.5B.3 at QTL10; Q.STB.6A.1, Q.STB.6A.3 at MQTL20; and Q.STB.7D.5 at MQTL26 region of a previously published QTLs [19]. The QTNs, Q.STB.1B.2 and Q.STB.1B3 on chromosome 1B were found close to SSR wmc206, which was linked to MQTL3. Similarly, QTNs, Q.STB.2A.6 and Q.STB.2A.7 were close (3-4Mb) to marker gwm294, conferring MQTL5 for STB resistance. The Q.STB_DH2A.1 was located close to MQTL4 and is responsible for adult plant resistance [19]. On chromosome 5A, we detected QTNs, Q.STB.5A.5 positioned at 417.22Mb and Q.STB_DH.5A.3 positioned at 588.45Mb, which are in the region of a previously published MQTL19 and QTL9 [3]respectively, conferring resistance to STB. However, both QTL9 and MQTL19 provide STB resistance at the seedling stage, while in our study, the QTNs were associated with adult plant resistance. Moreover, the QTN, Q.STB_DH.5A.3 also coincided with the Stb17 gene having a quantitative effect on disease at the adult plants stage [31]. Stb10 in chromosome 1D is a qualitative resistance gene imparting durable resistance to STB [32]. The QTL10 and MQTL9 are responsible for seedling resistance, while MQTL14, MQTL20 and MQTL19 are effective at seedling and adult plant stages. Interestingly four SNPs synchronizing with QTLs: MQTL3 at 1B chromosome, MQTL5 and MQTL4 on 2A chromosome and MQTL26 at 7D chromosome imparting adult plant resistance were also mapped. Brown et al. [19] also reported the involvement of QTLs on chromosomes 1B, 2A and 7D, imparting quantitative resistance to STB, which is true in our findings also. Hence, strategic use of the identified major and minor genes/QTNs can be used to deploy STB-resistant varieties.
Q.STB.1A.1, Q.STB_DH 2A.1, Q.STB.2A.3, Q.STB.2B.4, Q.STB.5A.1, Q.STB.5A.2 and Q.STB.7B.2 didn't coincide the location on the previously reported STB resistance loci in respective chromosomes. At the 7B chromosome, the Q.STB.7B.2 didn't coincide with Stb13, Stb3 or Stb 8 genes and any of the previously reported QTLs (MQTL25, MQTL26). Similar was the case with other QTNs. We have found novel QTNs on chromosomes 1A, 2A, 2B,5A and 7B which can be a good candidate for MAS for the STB resistance wheat breeding programme.
Examining the putative CG associated with significant SNP markers and STB resistance is crucial. Q.STB_DH 2A.1, 2A.3, 2A.6, and 2A.7 were mapped on chromosome 2A. SNP 3955868 at 106.81 Mb is associated with the putative candidate gene linked to this marker, TraesCS2A02G159300 (glycoside hydrolase superfamily). The loci covered glycosyl group manipulation genes. Molecular modifications by glycosylation change protein properties, activity, and target location. Glycosylation of metabolites and hormones occurs during biotic and abiotic stress. He et al. [33] found that wheat Glycosyl transferases convert Fusarium graminearum toxin DON into non-toxic DON-3-glucoside. QTNs Q.STB.2A.3 encodes CG TraesCS2A02G307700, Q.STB.2A.6 encodes TraesCS2A02G561400 and TraesCS2A02G561300, and Q.STB.2A.7 encodes TraesCS2A02G563700 and TraesCS2A02G563900, which are associated with metal ion binding, zinc ion binding, serine/threonine kinase activity. These proteins aid plant-pathogen interactions [34]. Saintenac et al. [11] cloned a major STB resistance gene (Stb6) that also encodes a conserved wall-associated receptor kinase-like protein. In wheat, inactivating serine/threonine kinase gene TaPsIPK1 confers broad-spectrum resistance to Chinese Pst races endemic in 2020 and 2021 [35]. Additionally, we found that chromosome 5A harboured protein binding, protein-coding and protein kinase domain (Peak marker 995502 of Q.STB.5A.1; 1228444of Q.STB.5A.2; 1023146 of Q.STB.5A.5 and Q.STB_DH.5A.3 (5411867). In wheat, Wang et al. [35] reported that the inactivation of a wheat protein kinase gene, TaPsIPK1 confers broad-spectrum resistance to rust fungi.
Further, TraesCS3B02G440700 (Q.STB_DH.3B.2) encodes protein kinase activity, while TraesCS3B02G442100 encodes aldose 1-epimerase and TraesCS3B02G442400 encodes cysteine-kinase. The major STB resistance gene Stb16q encodes a cysteine-rich receptor-like kinase and resists Z. tritici broadly [14]. Most R genes already identified have the ATPase domain, which is involved in biotic stress response and found in our CGs. The present study outlined the CGs encoded proteins, most of which confer plant disease resistance (Table 3).
Overlapping regions for STB resistance and disease -escape traits
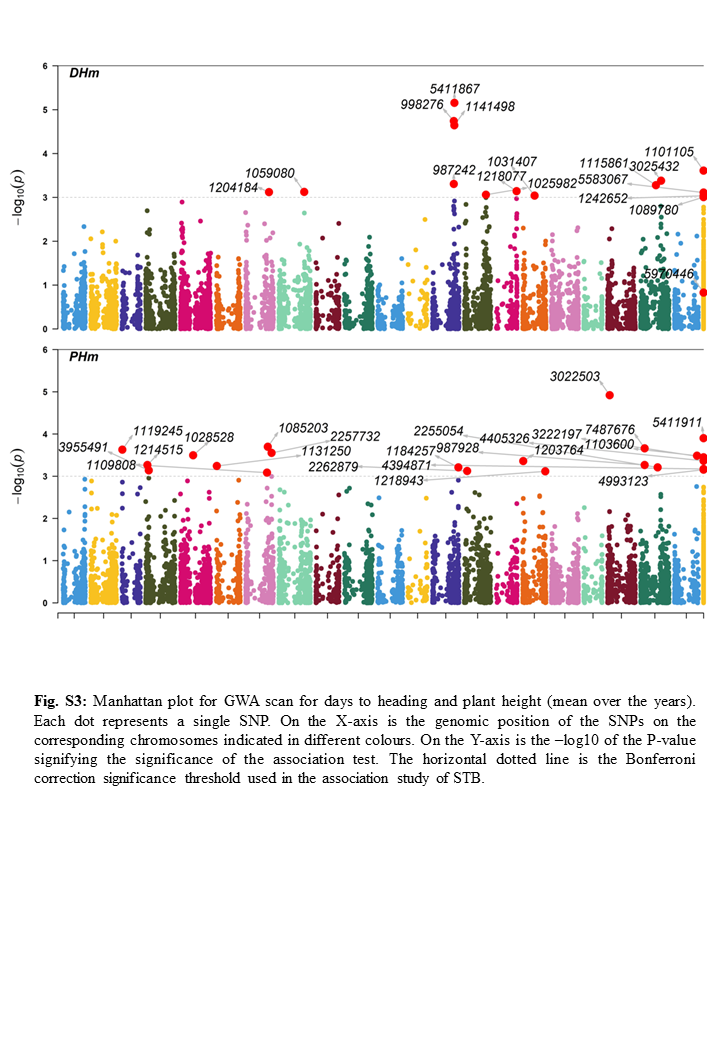
One of the most perplexing factors affecting the selection for STB resistance could be the reported interaction between resistance and plant height (PH) or heading date (DH) [36,37]. In the present study, a few loci were identified to be associated with both STB and DH on chromosome 2A, 3B, 5A, 5D, and unknown location Q.STB_DH.UN.5. Louriki et al. [30] also reported QTLs for heading days in wheat on chromosome 2A. Previous studies confirmed chromosome 5 for flowering and the presence of the vernalization gene, VRN-1 having three homoeologous loci VRN-A1, VRN-B1, and VRN-D1, which are reported on the long arm of homologous chromosomes 5A, 5B, and 5D, respectively [38,39]. The identified QTN in our study at 5A was close to VRN-A1. Also, for flower induction, the photoperiod response in wheat is mainly controlled by the PHOTOPERIOD1 (PPD1) loci located on the short arms of chromosomes 2A, 2B, and 2D. PPD1 genes identified in wheat are members of the pseudo-response regulator family [40]. The fact that in our panel, days to heading provide putative chromosome locations with QTNs for disease resistance at 2A, 5A and 5D chromosomes hypothesizes the presence of a common genetic base for plant structure, phenology, and disease susceptibility in the studied material. The present study also revealed that some of the significant QTNs for DH, co-mapped with previously identified STB-resistant regions like MQTL4 (Q.STB_DH.2A.1/3955868), MQTL14 (Q.STB_DH.3B.2/1059080), QTL9 and Stb17 gene (Q.STB_DH.5A.3/5411867) [19] thereby showed strong pleiotropism. QTN such as Q.STB_DH.3B.2/1059080 located near transcript TraesCS3B02G440800 related to magnesium ion transmembrane transporter activity has been reported to play a significant role in powdery mildew and stripe rust resistance in wheat [41]. The putative-associated regions identified in the present study need further study for confounding indirect selection for STB resistance via late heading.


{kind=link}
{kind=link}
{kind=link}