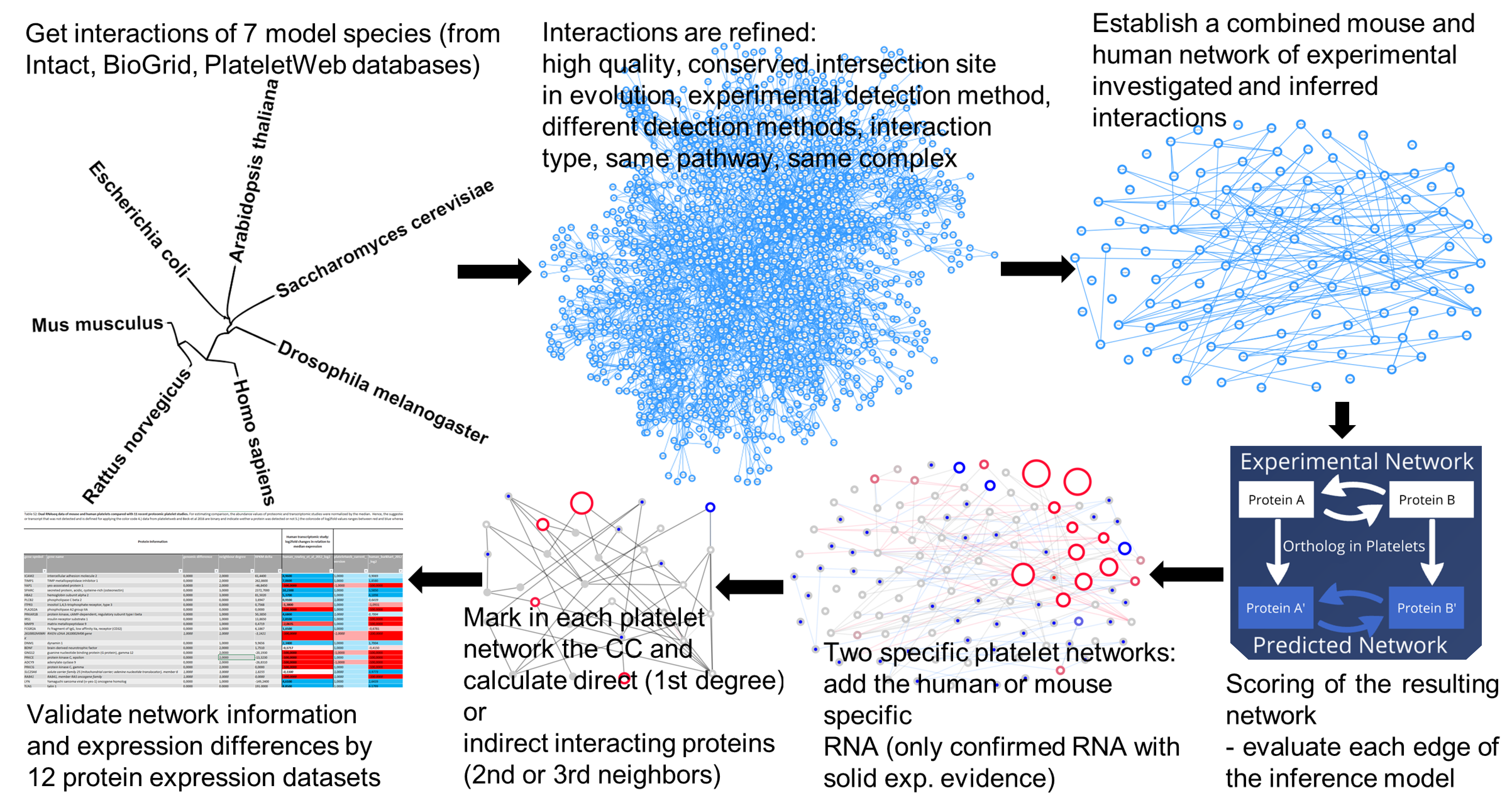
Using a recently published model of the central activating cascade of the platelet (CC; includes inhibition by cAMP) [19], we systematically compared the most important signaling cascades described in both human and mouse platelets. For this study, we have used integrated genomic data followed by complete ortholog mapping of transcriptome and proteome datasets to compare the CC between mouse and human, using only correctly corresponding proteins and genes (orthology) and test their expression levels using platelet RNA and protein datasets. All available platelet transcriptome data were used to screen and evaluate potential differences between human and mouse. For meticulous validation, we used eleven recent high-quality proteome and phosphoproteome datasets [20-30] and compared them (detailed information in Supplemental Material and Methods). To consider only validated protein-protein interactions, we mapped our large-scale genome/transcriptome and proteome datasets using a reconstructed protein interactome of mouse and human platelets (see methods and Table S1 where full protein names are given). We next considered all available further large-scale and specific experimental data to validate differences between mouse and human (Table S2; full names included). In 46% of the cases, we could confirm RNA expression differences by similar protein expression differences after normalization (Table S2). In further 35% of the cases, the evidence was only compatible with the prediction, the available information from the proteome was not conflicting with the observed RNA difference (Table S2). It is important to note that in 19% of the cases proteome and RNA expression data showed clear opposite differences between the species (Table S2), suggesting different regulation on RNA or protein level and requiring further experimental investigation. This concerned four proteins (SRC, TBXA2R, PTGDR, RASGRP1) in the central cascade, as well as 8/37 1st neighbors plus 4/38 2nd degree neighbors (proteins explained in Supplemental File 1, data in Table S2; 99 mRNA differences investigated).
In the next analysis step, we focused on all confirmed expression differences. The combined data compared the same proteins in mouse and human (direct seed orthologs) to reveal differences (Figure 1; detailed full network in Figure 2). In total, 1132 proteins were confirmed to have the same function in both species (all are direct orthologs). Table 1 lists the species networks for human and mouse. 621 human mRNAs are solely contained in the human network and 58 murine mRNAs are only found in the murine network. Besides species-specific variation in proteins found in human and mouse platelets, this results in species-specific subnetworks including differences for well-connected orthologs (same protein in mouse and human with more or less or sometimes different connections depending on species). The proteins in the networks are represented according to their mRNA evidence.
Using the similarity of conserved pathways the combined network supports the network reconstruction of each species. The current reconstructed network of human platelets encompasses 1608 proteins and 9406 interactions (Figure S2). The murine network comprises of 1051 proteins and 5769 interactions (Figure S3). The direct comparison of each species network covers 858 direct ortholog proteins and 3648 shared interactions. The combined network (Figure 2) has 1801 proteins. Half of these proteins (903) are abundant in platelets in at least one of the two species (RPKM > 3; adjusted threshold according to the median of the central cascade).
Key results (asterisk) of this comparison between mouse and human are indicated in Figure 1 and summarized in Figure 3, individual differences are discussed in Supplemental Material taking all available proteome and RNA datasets into account.
Overall expression and network differences
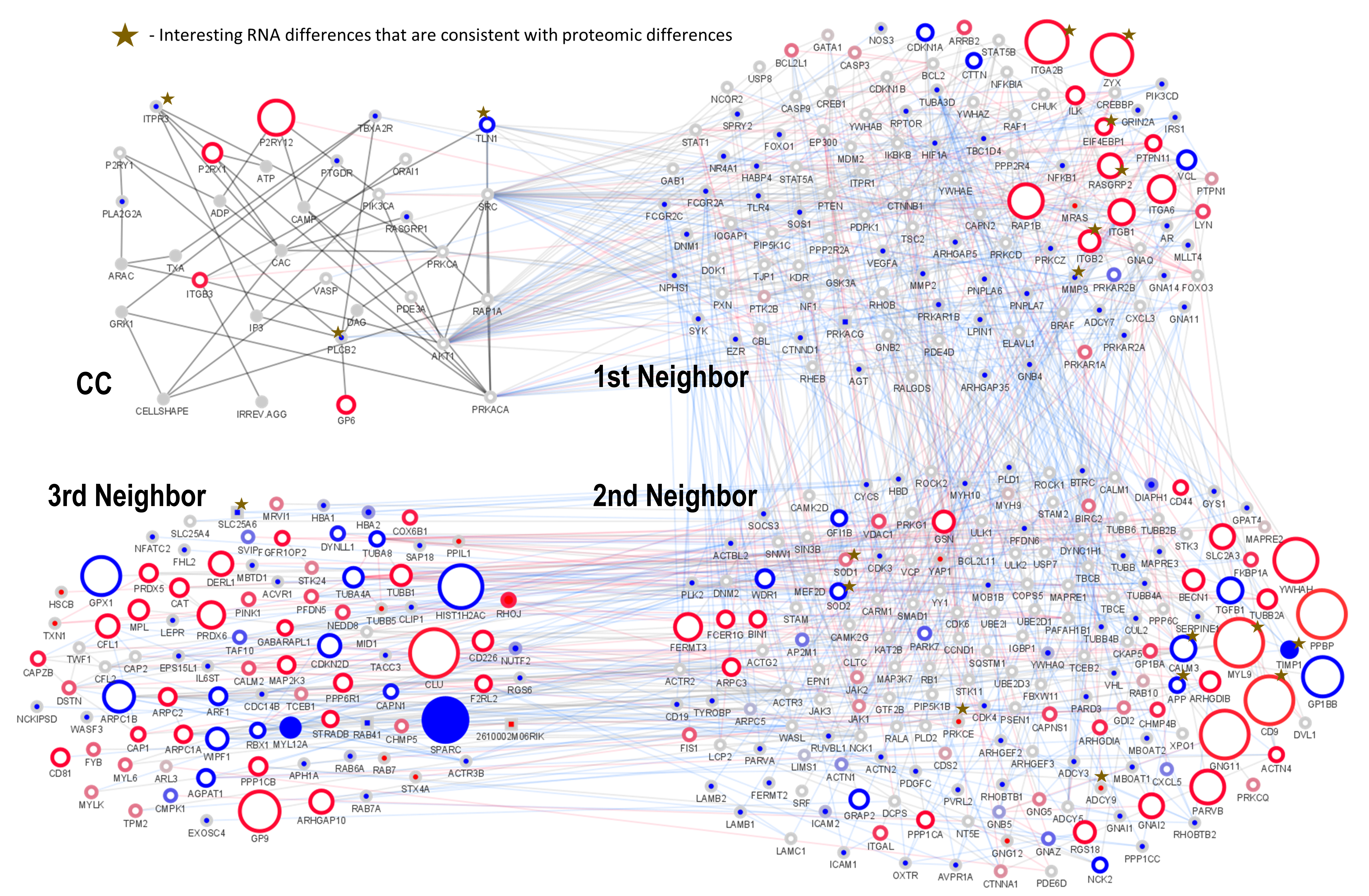
The overview of the central regulatory proteins and the central cascade shows that murine proteins involved in platelet signaling are expressed at higher levels (median RPKM: 4.5) compared to human platelets (median RPKM: 2). The cumulative expression (RPKM) in mouse was also much higher (total RPKM: 96420) compared to human (total RPKM: 53487). We found that the well-studied human signaling network includes a higher number of proteins (1608) compared to model organism mouse (1051). In the human network up to degree 3, we identified 33 proteins with a relatively high RPKM (over 100). In contrast, within the mouse signaling network, 82 proteins were detected with high RPKM (more than 100). The full central network with all regulators up to neighbor degree 3 results in 1618 nodes and 9406 edges in human, and 1061 nodes and 5769 edges in mouse (including non-protein interactors). Our calculation included also the signaling molecules which belong to the CC, according to Mischnik et al., [19] Although the CC is assumed to be conserved between mouse and human species, using all available information from databases and experiments, we found a number of clear genetic differences as well as different mRNA and protein levels in mouse and human platelets. Figure 1 shows the resulting network (asterisks label key differences), the CC and its neighbors, including 369 nodes and 1646 edges in human and 277 nodes and 1119 edges in mouse.
In addition, we also compared the total platelet network of mouse and human (Figure 2). The human network contains 3524 nodes with 18618 high confidence protein interactions (almost certainty; p > 0.99). The average number of protein interactions was about 5 interactors per signaling protein. In comparison, a high confidence dataset in the IntAct database [31] reports 9 interactors per protein and only 6 interactors by excluding high binders. In sharp contrast, the complexity of the mouse network was found to be reduced, only 2114 nodes and 10337 interactions were identified. Nevertheless, similar network properties were found and the average number of interactors was 5 per protein. The overall analysis presented here has no species bias using a homogenous prediction method. All major differences found for the CC, its direct neightbors and 2nd or 3rd degree neighbors are concisely summarized in Figure 3.
Specific differences in the central cascade
The systems biological defined CC [12, 32, 33] showed no genomic difference between human and mouse platelets (Figure 3). However, abundance differences of mRNA and protein could be identified in the CC (Fig. 1; blue borders indicate higher expression in human and red indicates higher expression in mouse; proteins directly interacting with the CC are 1st degree neighbors of the CC, interactors of these are 2nd degree neighbors and the proteins interacting only with the 2nd degree neighbors in turn are 3rd degree neighbors). PLCB2 (phospholipase C beta 2) and ITPR3 have not been detected in mouse on mRNA level, but are expressed in human (matches proteome evidence). Talin (TLN1) mRNA is higher abundant in human which is confirmed by proteomics (Table 2).
RNAseq and proteome datasets could not provide firm evidence for the detection of relevant expression levels of Phospholipase A2 Group IIA (PLA2G2A) in both species in transcriptome, as well as proteome. TBXA2R (thromboxane A2 receptor) shows a higher protein level in mouse but the absence of mRNA in mouse and high mRNA expression in human. PTGDR (prostaglandin D2 receptor (DP)) only has mRNA expression in human and no protein evidence was found in both species. The same is valid for RAS guanyl-releasing protein 1 (RASGRP1). Purinergic receptor signaling is regulated by P2RY12 (purinergic receptor P2Y, G-protein coupled, 12) and P2RX1 (purinergic receptor P2X, ligand-gated ion channel 1). mRNA expression levels of these receptors, which are directly activated by ADP and ATP, respectively, [34, 35] are clearly higher in mouse. In accordance with this, there is clear protein quantification of P2RY12 receptor in murine platelets (log2: 1.3; Zeiler et al., 2014 [27] and 2.0 according to Hurtado et al., 2018 [28]) but P2RY12 protein in human is low and difficult to detect (Table S2). It is present in really low and variable amounts [36] but easy measured as functionally present receptor [37]. These concordant results of mRNA and proteome support a difference in central receptor signaling between mouse (higher expression of P2RY12) and human. For the calcium channel P2RX1 and the collagen receptor GP6 (GPVI in human platelet) higher mRNA expression in mouse was found but proteome data suggest opposite protein abundance. ITGB3 (integrin beta 3) differs slightly on mRNA level, but not on protein level. In addition, the highly expressed central platelet signaling kinase SRC [10] shows clear differences, although mRNA and protein level give opposite estimates suggesting independent regulation.
Major mouse-human platelet proteome expression differences
Key proteome differences of the CC are summarized in Table 1. There are clear differences in the regulation and modulation of the central cascade between man and mouse. In particular, sometimes a protein counter part in the other organism is lacking or almost absent, there are strong expression differences. Each of these clear differences with functional implications for the platelet has been several times reported and observed in literature (Table 1).
Higher abundance of copper-zinc-superoxide dismutase 1 (SOD1) in murine platelets implies better ROS protection [38]. In human platelets, manganese-dependent superoxide dismutase 2 (SOD2 in the mitochondrial matrix) is higher abundant (Table S2). It regulates apoptotic pathways and expression differences influence also platelet apoptosis-like activation [39].
The talin abundance difference is important as it regulates key proteins in platelets [11] such as integrin influencing thrombosis and platelet adhesion[40]. In particular, Talin decreases integrin activation and reduces the probability of the platelet for irreversible aggregation [41].
Glycoprotein VI (GP6 or GPVI), the platelet receptor for collagen, laminin and fibrin, centrally regulates multiple platelet functions, including adhesion, activation, aggregation and pro-coagulant activity [42-47].
Matrix metalloproteinases (MMPs) are reorganizing the extracellular matrix [48, 49]. MMP9 is only present in humans, its low expression affects platelet activation [50] (Table S2 and [51]). Platelet expression differences in tissue inhibitors of metalloproteinase (TIMPs), such as TIMP1, TIMP2 and TIMP3 affect activities of MMPs and by this platelet aggregation [52, 53].
Brain-derived neurotrophic factor (BDNF) is described only in humans. Protective effects for brain [54] are mediated by platelet BDNF and impaired by smoking [55, 56]. There are gender differences as well as BDNF expression differences in patients with cardiovascular disease and depression. All these brain protective effects mediated by platelet BDNF are absent in mouse platelets, there is no similar protein (ortholog) present. Src protein kinase shows opposite differences regarding mRNA versus protein level regulation in man and mouse (Table S2). This implies that Src kinase as the central bistability switch of the activating cascade [33] has different activation tipping points in man and mouse.
More detailed functional relevance of proteins pointed out in the paper or found to be different in the CC network are given in the supplementary material.
Specific genomic differences in the 1st to 3rd degree neighbors
Five human genes not detected in the mouse genome are 1st to 3rd degree neighbors of the central cascade: SLC25A6, CASP10, PRKACG, HSPA6 and RAB41. Details are given in Table S2 and the Supplemental data file considering all available data-sets.
Different expression profile of the 1st to 3rd degree neighbors
We looked at proteins directly interacting with the central cascade (1st neighbors of CC) or interactors of these (2nd neighbors of CC) or one interaction further (3rd degree neighbors of CC) using well established human and murine interaction data. A first screen analyzed mRNA expression differences in both species after normalization, next a detailed comparison according to support or lack of support in the eleven large-scale platelet proteome studies was done (see materials and methods for details including log2 value calculations and comparison protocols).
According to this census of all available data, the following further differences were found for direct interacting protein neighbors (1st neighbors) of the central cascade analyzing their differential expression:
There are 44 proteins, which are not detected in mouse, but identified in human platelets. Within this group, PRKAR1B (log2: 4.7 (mRNA) and 0.7 (protein)), IRS1 (log2: 2.8 (mRNA) and -0.4 (protein)), DNM1 (log2: 2.3 (mRNA) and 2.7 (protein)) and FCGR2A (log2: 1.7 (mRNA) and 0.5 (protein)) are the most relevant. HABP4 has only mRNA but no protein expression in human (log2: 1.5 for mRNA; no detection in any of the proteome studies), thus suggesting the expression of a non-coding RNA (ncRNA); XR_001746249; miscRNA). MRAS and KDR are clearly detected as murine mRNA (log2: 3.2 and log2: -0.6) but lack protein evidence in both species. Similar, mouse mRNAs of DOCK1, NF1 and TJP1 were only detected in mouse but protein levels are unclear, or in the opposite expression difference to the mRNA expression level differences. DOCK1 is slightly expressed on protein level with a small tendency to be increased in murine mRNA and protein. NF1 and TJP1 levels are low when present, indicating the proximity to detection sensitivity limits. A higher mRNA expression level of four proteins, namely VCL, CDKN1A, CTTN and Protein Kinase CAMP-Dependent Type II Regulatory Subunit Beta (PRKAR2B) was found in human platelets. Proteome datasets supported these differences for VCL and CDKN1A, although the extent is lower on protein levels. For CTTN high abundance levels are detected in transcriptomics, as well as in proteomics for mouse and human, with a trend for higher levels in humans. PRKAR2B is similarly confirmed to be an abundant protein in both species, but shows clearly higher protein level in mouse.
In mouse platelets, 17 mRNAs were found to be expressed higher than in human platelets, such as Integrin αIIβ, RAP1B (matching several proteome data-sets), ITGA6 (proteome data find the opposite species difference to the RNA data), ITGB1 (compatible), RASGRP2, ITGB2, ZYX, ILK (matches proteome data), EIF4EBP1, PTPN11 (matches), LYN (matches), ARRB2, PRKAR1A (compatible), BCL2L1 (compatible), CASP3 (matches), PTPN1 (matches), PTK2B (matches) and STIM1 (matches proteome; full information and names are given in Table S1, S2). RASGRP2, ITGB2, EIF4EBP1, ARRB2, and ZYX are particularly interesting candidates for further investigation with substantial mRNA differences that are in accordance with proteomic differences.
Regarding the 2nd neighbors (543 human mRNAs) we identified 134 proteins in human such as TIMP1 which were not found in mouse platelets. There are 46 direct ortholog mRNAs that are clearly higher abundant in mice such as CD9 and SLC2A3 though also detected in human (details in Supplementary Data).
Referring to mRNA levels, in the 3rd degree neighbor network, 285 mRNAs are only detected in human, but not in mouse platelets, whereas 58 mRNAs are exclusively detected in mouse platelets. 24 of the 3rd neighbor mRNAs show a stronger expression in human. 46 platelet mRNAs
{kind=link}
{kind=link}
{kind=link}
{kind=link}