Morphological analysis
Of the three leaf characters, leaf length (L), leaf width (W) and ratio of leaf length to width(L/W), putative hybrid individuals consistently had two morphological characters, leaf length and leaf width, intermediate in value between B. alternifolia and B. crispa (Table 1). The L/W were significantly larger in B. alternifolia than the other two taxa (Table 1). Of the seven floral characters, corolla tube width (TW) and anther height (AH) of B. × wardii were intermediate between the values of the two assumed parental species, whereas herkogamy (HE) did not differ significantly between the three taxa (Table 1). The remaining four floral characters, corolla tube length (TL), corolla lobe length (CLL), corolla lobe width (CLW) and style length (SL) all showed a similar pattern, with the characters in B. alternifolia having significantly lower values than those measured in B. crispa or B. × wardii (Table 1).
The two putative parental species are morphologically clearly distinct. In the PCA of 10 morphological characteristics, 52.17% and 12.98% of the variation in total was explained by the top two principal components, respectively. The two-dimensional scatter diagram based on PC1 and PC2 showed clearly the separation of B. alternifolia and B. crispa. Individuals of B. × wardii fell between the two parent species, with a slight overlap with B. alternifolia and a large overlap with B. crispa. Apart from the character HE, there is little difference in the correlation coefficients between the other nine traits (0.29-0.38) (Figure 2a).
Petal color reflectance
Difference in reflectance spectrum of corolla was showed among there three taxa. Both B. crispa and B. × wardii were observed at 485 nm with obvious peak in the reflectance spectrum, with extremely low variation. However, there was no obvious peak in the reflectance spectrum in B. alternifolia (Figure 2b).
Pollination treatments
These two interspecific cross-pollination treatments with B. crispa or B. alternifolia as the mother species showed significant differences in fruit set, mean number of seeds per fruit, seed set, and mean number of vigor seeds per fruit. When B. crispa is the maternal plant, higher fruit set (64.71% vs 6.45%), more seed numbers per capsule (43.42±16.01 vs 6.5±0.71), higher seed set (68.79% vs 0) and more vigor seeds per fruit (26.90 vs 0) were coincidentally examined when comparing these parameters to cross-specific pollination when B. alternifolia as maternal plant (Supplemental Table S1). Additionally, comparatively large amount of seeds was produced when B. crispa was attributing to geitonogamy pollination indicating self-compatible breeding system (seed set: 48.68%) whereas no seeds were obtained when B. alternifolia was attributing to hand self-pollination.
Sequence analyses of the four nuclear genes
NrETS: The total length of the nrETS region alignment was 380 bp in all individuals, including 30 nucleotide substitutions and one 1-bp insertion/deletion (for variation sites, Supplemental Table S2). These variable sites generated 40 haplotypes, among them five, six and thirteen haplotypes belong to B. alternifolia, B. crispa and B. × wardii in BH population, whereas six, three and seven haplotypes in TJ populations, respectively. For the four reference populations, samples of JZA, KDA, BSC and KMC had two, three, three and four specific haplotypes, respectively. Two major clusters were identified by haplotype network analysis with six nucleotide mutations. One cluster composed mainly haplotypes of B. alternifolia and B. × wardii, comprising five haplotypes of BHA, one haplotype of BHC, seven haplotypes of BHW, six haplotypes of TJA, one haplotype of TJC, three haplotypes of TJW and all haplotypes of B. alternifolia from allopatric populations. The other cluster was mainly consisting of haplotypes of B. crispa and B. × wardii, including five haplotypes of BHC, six haplotypes of BHW, two haplotypes of TJC, four haplotypes of TJW and all haplotypes of B. crispa from allopatric populations (Figure 3 a).
Two haplotypes from B. crispa nested with the B. alternifolia cluster, and was found in BHCR2, BHCR7 and TJCR13, which show different haplotypes from both clusters. Except for two individuals, BHWI9 and TJWI1, all B. × wardii individuals with two haplotypes clustered into two divergent clusters and shared the mainly haplotypes of both B. alternifolia and B. crispa from the BH and TJ. The remaining two individuals had only one haplotype shared with B. crispa and were homozygous at this locus (Figure 3a).
GapC2: The total length of the gapC2 region alignment was 606 bp for all individuals, including 67 nucleotide substitutions, one 2-bp and one 1-bp insertion/deletion (for variation sites, Supplemental Table S2). A total of 41 haplotypes were observed from these loci among which one, nine and nine haplotypes belong to B. alternifolia, B. crispa and B. × wardii in BH population, whereas three, six and ten haplotypes in TJ populations, respectively. For the four reference populations, samples of JZA, KDA, BSC and KMC had one, five, eleven and four specific haplotypes, respectively. The haplotype network fall into three major clusters with one cluster consisted of all haplotypes of KDA, the second cluster contained one haplotype of BHA, one haplotype of BHC, two haplotypes of BHW, two haplotypes of TJA, three haplotypes of TJW, and the only one haplotype of JZA that share with the main haplotype of TJA. And the third cluster contained eight haplotypes of BHC, seven haplotypes of BHW, six haplotypes of TJC, seven haplotypes of TJW and all haplotypes of B. crispa from allopatric populations.
For B. crispa in both populations of TJ and BH, two individuals (BHCR2 and BHCR7) had a haplotype found in the second cluster but all other haplotypes were found in the third cluster. For B. alternifolia, there was an exception of two individuals, TJAL6 and TJAL12, which had a haplotype found in the third cluster, but all the other haplotypes were found in the second cluster. All B. × wardii individuals but one (BHWI9) showed two divergent haplotypes originating from both the second and third clusters and shared the mainly haplotypes of both B. alternifolia and B. crispa from the BH and TJ. The remaining individual BHWI9 was homozygous for a haplotype from the third cluster (Figure 3b).
PPR24: The total length of the PPR24 region alignment was 647 bp for all individuals, including 70 nucleotide substitutions (for variation sites, Supplemental Table S2). This locus has the maximum haplotypes of 53. B. alternifolia, B. crispa and B. × wardii in BH populations had four, ten and 14 haplotypes, whereas there were one, 11 and 15 haplotypes in TJ population, respectively. For the four reference populations, samples of JZA, KDA, BSC and KMC had one, one, seven and seven specific haplotypes, respectively. In the haplotypes network analysis, two major clusters were identified with 16 nucleotide substitutions. One cluster contained four haplotypes of BHA, one haplotype of BHC, seven haplotypes of BHW, the only haplotype of TJA, eight haplotypes of TJW and the unique haplotype for each of the two referenced populations of B. alternifolia. The other cluster contained nine haplotypes of BHC, seven haplotypes of BHW, all haplotypes of TJC, seven haplotypes of TJW, and all haplotypes of B. crispa from allopatric populations. One B. crispa individual (BHCR7) and all B. × wardii individuals showed two divergent haplotypes originating from both clusters (Figure 3c).
PPR123: After sequence alignment, the PPR123 region was 735 bp in length for all samples including 59 nucleotide substitutions (for variation sites, Supplemental Table S2). A total of 45 haplotypes were identified for this gene, among which two, four and eight belong to B. alternifolia, B. crispa and B. × wardii in BH population, whereas two, ten and ten haplotypes in TJ population, respectively. For the four reference populations, samples of JZA, KDA, BSC and KMC had two, four, eleven and two specific haplotypes, respectively. As shown in the haplotypes network, all haplotypes were clustered into four groups with seven, six or 13 variations between them. The first clade comprised two haplotypes of BHA, one haplotype of BHC, five haplotypes of BHW, two haplotypes of TJA, one haplotype of TJA and all haplotypes of B. alternifolia from allopatric populations. The second clade comprised three haplotypes of BHC, three haplotypes of BHW, five haplotypes of TJC and seven haplotypes of TJW. The third clade comprised five haplotypes of TJC and two haplotypes of TJW. The remaining clade comprised all haplotypes of B. crispa from allopatric populations (Figure 3d).
All haplotypes from B. alternifolia individuals fell into the first cluster. For the B. crispa individuals in BHC and TJC, only one from BHC nested with the first clade derived from BHCR7, ten individuals (TJCR3/5/6/7/9/10/11/12/13/15) had two divergent haplotypes, falling into the second and third clusters, four individuals (TJCR1/4/8/16) had haplotypes nested in the second clade and two individuals (TJCR2/14) had haplotypes nested in the third clade. Of individuals of B. × wardii, all but BHWI9 had two different haplotypes, one clustered in the first cluster and share the major haplotypes with B. alternifolia, the other clustered in either the second or the third cluster and share the major haplotypes with B. crispa. The remaining BHWI9 had two haplotypes found in the second clade (Figure 3d).
Sequence analyses for the combined chloroplast regions
The combined aligned length of the cpDNA fragment alignment (including rpl16, trnD-trnT, trnS-trnfM) was 2014 bp, and contained 38 nucleotide substitutions, one 1-bp, one 2-bp and one 6-bp insertion/deletion (for variation sites, Supplemental Table S2). A total of 15 haplotypes were found in all samples, among which three, three and one were specific to the reference population of KMC, BSC and KDA, respectively. Haplotype network analysis indicated that these three reference populations differed from other populations by at least seven DNA base variants. All individuals in TJ had only two haplotypes, of them one was derived from six individuals of TJA (TJAL2/3/4/5/8/11) and shared with the only haplotype from the reference population JZA, and the remaining 42 individuals shared another haplotype consistent with the haplotype only in individual BHAL13. Each taxon had three haplotypes in BH population, among which two, one and one were specific to B. alternifolia, B. crispa and B. × wardii, respectively. Most of the individuals in BHW (75%) shared haplotypes with B. crispa (BHC), while they did not share any haplotypes with B. alternifolia. The remaining four B. × wardii individuals (25%) had a unique haplotype with one mutational step away from the predominant haplotype of B. crispa (Figure 3e).
NewHybrids analysis
Analysis of the four studied nuclear genes using NewHybrids among the three taxa showed that individuals morphologically identified as B. × wardii in TJ and most individuals in BH were F1 hybrids between B. alternifolia and B. crispa. At BH, 31 of 33 individuals with putative parental morphology were assigned to be the pure parental species with high posterior probabilities (>98.8%). All individuals of BHW but two (BHWI2 and BHWI9) were identified as F1s with >96.8% posterior probability. Of these two, BHWI2 was identified as F1 with a 77.8% posterior probability, whereas BHWI9 was identified as B. crispa with a 99.7% posterior probability (Figure 4a).
At TJ population, only TJCR13 was identified as B. crispa with low probability of 84.0%, and all the rest of the putative parental individuals had a high posterior probability of >99.0% to be recognized as pure parents. The posterior probabilities of all putative hybrids being presumed to be F1s were >97.4% (Figure 4a). Therefore, in both populations, three individuals were identified as backcross to B. crispa with a probability of less than 16%, which was far lower than their probability of being B. crispa or F1s, so the hybridization was not beyond the F1s.
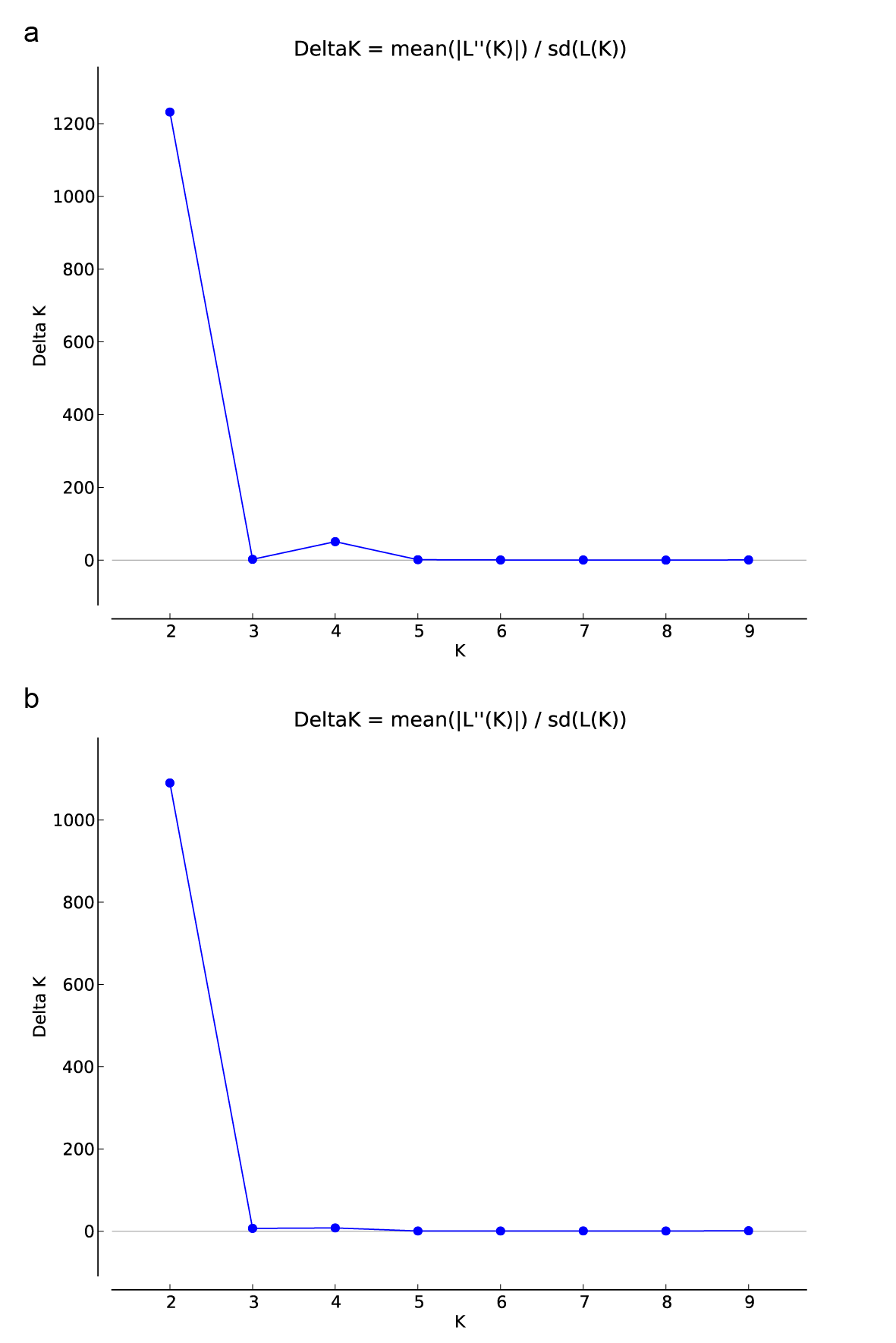
Population structure analysis
Structure analysis of a total of 213 variation loci for all individuals showed that the highest value of △K was obtained when K=2, suggesting that all 152 individuals were clustered into two types of genetic clusters. When K=2, for the four reference allopatric populations, all individuals of KMC and BSC formed a pure cluster (q≥0.999), while all individuals of KDA and JZA formed another pure cluster (q≥0.998).
As showed in the Figure 4b, in BH population, all morphologically determined to be B. alternifolia clustered into one cluster with their high proportion containing one genetic component (q≥0.997). Except for two individuals, BHCR2 and BHCR7, all individuals identified as B. crispa formed another cluster with the proportion containing another genetic component ≥0.998. While BHCR2 and BHCR7 contained the same genetic component of BHC were 0.755 and 0.510, respectively. All B. × wardii individuals showed approximately equal proportions from both clusters (q=0.494±0.084 for BHA), expect for BHWI9, which had a proportion of 0.820 genetic component from BHC cluster.
Similar to the BH population, most of the B. alternifolia (TJA) and B. crispa (TJC) in the TJ population were segregated into a high proportion of different genetic components (q≥0.988 for TJA; q≥0.999 for TJC). Of the remaining three individuals, the probability that TJAL6 and TJAL12 contained low levels of genetic components from TJA was 0.959 and 0.931, respectively, while TJCR13 possessed genetic components from TJC was 0.830. All individuals of B. × wardii (TJW) possessed close proportions of genetic components from both TJA and TJC (q=0.523±0.033 for TJA).
{kind=link}