PROS1 is not expressed by myeloma plasma cells but is enriched in the bone marrow of MM patients
To explore the role of PROS1 in MM, we analyzed PROS1 mRNA expression by Affymetrix array in sorted (FACS) CD138+ bone marrow plasma cells (BMPCs) from 332 newly diagnosed MM patients in comparison to blood mononuclear cells (MBC) and BMPC isolated from healthy donors and monoclonal gammopathy of unknown significance (MGUS) patients. The analysis revealed that PROS1 was not expressed by the majority of BMPCs from MM and MGUS patients as well as healthy controls at the mRNA level (Fig. 1A).
Various myeloma-promoting cytokines and growth factors are secreted by the stroma. We measured PROS1 protein by ELISA in BM plasma samples of newly diagnosed MM patients in comparison to healthy donors and detected increased PROS1 levels in BM plasma of MM patients (Fig. 1B). Analysis of PROS1 mRNA expression in BM cells of MM patients in comparison to healthy controls revealed slightly increased PROS1 expression, but results did not reach statistical significance, which may reflect the fact that MM plasma cells mainly do not express PROS1 (Fig. 1C). Together, these results suggest that PROS1 might be upregulated by the tumor microenvironment in the BM of MM patients, whereas MM cells mostly do not secrete PROS1.
PROS1 regulates MM plasma cell growth in vitro depending on its concentration.
To investigate a potential role of PROS1 in MM plasma cells, we expressed PROS1 in MM cell line U266 using lentiviral gene transfer (Suppl. Fig.1A). Overexpression of PROS1 in U266 cells resulted in increased growth rates in baseline (Fig. 2A). In concordance with increased growth upon PROS1 overexpression, treatment of MM cells with PROS1 purified from human plasma led to increased proliferation of U266 cells and GAS6-dependent cell line RPMI8226 upon subnanomolar concentrations of PROS1 in a dose-dependent manner (Fig. 2B and C). These concentrations were in line with PROS1 concentrations measured in cell culture supernatants (100 ng/ml = 1.3 nM) (Suppl. Fig. 2). Similar results were obtained by treating U266 and RPMI8226 cells with recombinant GAS6 in subnanomolar concentrations, suggesting that PROS1 can induce MM growth comparably with GAS6 in vitro (Fig. 2D and E). The subnanomolar dose ranges stimulating MM growth in vitro correspond to human plasma levels of GAS611. However, previous data indicate that human plasma levels of PROS1 are around 1000-fold higher than those of GAS618. Therefore, we treated MM cells also with higher doses up to 100 nM PROS1. Interestingly, high PROS1 concentrations led to growth inhibition in U266 and RPMI8226 cells, suggesting that high PROS1 levels in the environment are detrimental for MM plasma cells (Fig. 2F and G). In addition, we observed increased chemosensitivity towards proteasome inhibitor Bortezomib (BTZ) upon overexpression of PROS1 (Fig. 2H).
PROS1 inhibits MM progression in vivo
In a next step, we injected U266 cells with transgenic expression of PROS1 intrafemorally into NSG mice. λ light chain concentration was measured as a surrogate marker for tumor load. Interestingly, we observed decreased λ light chain concentration in mice injected intrafemorally with U266 cells harboring PROS1 overexpression at week 9 (Fig. 3A). Furthermore, these mice showed prolonged overall survival, indicating that high PROS1 environments inhibit MM progression in vivo (Fig. 3B). We measured bone marrow plasma cell infiltration by flow cytometry (FC) in the contralateral leg and observed decreased MM cell numbers, suggesting that increased PROS1 levels might inhibit MM dissemination by inducing bone marrow retention (Fig. 3C and D).
PROS1 induces an adhesive and quiescent MM plasma cell phenotype via AXL
MM progression is followed by dissemination processes through bone marrow niches and in advanced stages to extramedullary sites19. MM cells can develop invasive properties relying on cell adhesion molecules and chemokine receptors20-22.
TAM receptors regulate cellular adhesion and migration23. We hypothesized that PROS1 could alter adhesive interactions of MM cells with the microenvironment, processes controlling MM dissemination and progression. U266 is a semi-adherent cell line that grows in tissue culture flasks as a mixture of adherent and suspension cells24. We cultured U266 MM cells on plates coated with extracellular matrix (ECM) glycoprotein fibronectin and detected increased adherence of MM cells to fibronectin upon treatment with PROS1 (Fig. 3E). We performed immunofluorescence staining of filamentous actin (F-actin) in U266 MM cell cultures on fibronectin to characterize MM morphology. U266 MM cells usually appear round and contracted reflecting myeloma plasma cell phenotype. In contrast, treatment with PROS1 led to high numbers of MM cells with a flat and elongated morphology, suggesting that PROS1 regulates cell morphology and adhesion in MM plasma cells (Fig. 3F). Interestingly, PROS1 promoted spontaneous myeloma cell aggregates (Fig. 3F, white arrows). As we detected decreased MM progression upon PROS1 overexpression in an orthotopic in vivo model, we conclude these effects might be mediated by increased MM adhesion resulting in BM retention of MM PCs. Interestingly, in previous studies it was shown that AXL induces quiescence and dormancy in MM15. As PROS1 drives AXL expression in oral squamous cell carcinoma, we hypothesized that PROS1 might upregulate AXL to induce growth arrest in MM25. Indeed, we measured increased AXL expression induced by PROS1 in U266 cells, suggesting that PROS1 might induce an adhesive and quiescent MM phenotype via upregulation of AXL (Fig. 3G).
PROS1 is highly expressed by cells of the bone marrow microenvironment
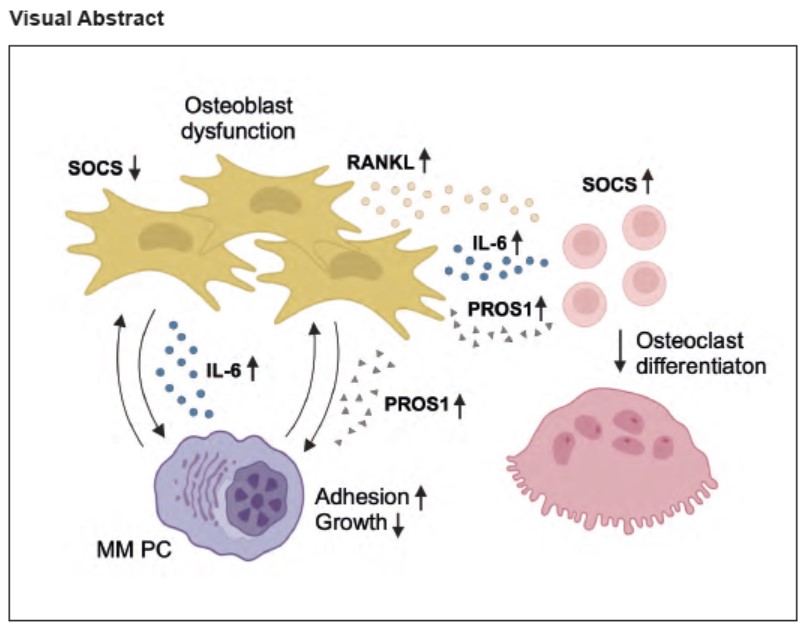
Since we measured increased PROS1 levels in BM plasma of MM patients but unraveled a rather negative-regulatory role in MM plasma cells, we next investigated its biological effects on the MM tumor microenvironment. The interaction of MM with BM niches favors myeloma migration, growth, survival and myeloma bone disease. The MM tumor niche comprises the components of the non-cellular (extracellular matrix) and cellular microenvironment compartment consisting of stromal cells, osteoblasts, osteoclasts, endothelial cells and immune cells4. To unravel the potential source of elevated PROS1 levels in MM BM microenvironment, we measured PROS1 expression in the different cellular compartments. We screened for expression in murine mesenchymal stem cell line OP-9, primary murine osteoblast cultures, endothelial cell line HUVEC, primary murine osteoclast cultures and in three different human myeloma cell lines U266, RPMI8226 and JJN3. We detected PROS1 expression in all stromal cell compartments and identified osteoblasts as the main cell type expressing PROS1. As expected, all myeloma cell lines showed either no or low PROS1 expression (Fig. 4A). We hypothesized that MM cells might induce expression of PROS1 in the osteoblast lineage, thereby causing increased PROS1 levels measured in BM samples of MM patients. Indeed, we detected increased Pros1 expression in osteoblasts when co-cultured with MM cell lines U266, RPMI8226 and JJN3 (Fig. 4B).
PROS1 – IL-6 crosstalk in MM plasma cells and osteoblasts
IL-6 is one of the main cytokines in MM promoting tumor progression, interaction with the BM microenvironment and fostering myeloma bone disease6. IL-6 and GAS6 are secreted by BM stromal cells and induce MM proliferation and survival. Additionally, IL-6 upregulates GAS6 expression and vice versa in MM cells and BM stromal cells, thereby inducing a vicious cycle11. In order to investigate a potential link between IL6 and PROS1, we treated U266, RPMI8226 and JJN3 MM cell lines with IL-6 and detected increased PROS1 expression, even in RPMI8226 cell line with undetectable PROS1 expression in baseline (Fig. 4C-E).
We hypothesized that MM cells might induce increased expression of PROS1 in osteoblasts via IL-6, but treatment with IL-6 did not show any effect on Pros1 expression in osteoblasts (Fig. 4F). Vice versa, osteoblasts treated with PROS1 showed interestingly highly increased Il6 expression levels (Fig. 4G). These results led us hypothesize that PROS1 induces proinflammatory signaling in osteoblasts.
PROS1 promotes IL-6 secretion by osteoblasts via inhibition of SOCS expression
To elucidate the underlying mechanism, we investigated PROS1-mediated downstream signaling pathways leading to increased IL-6 secretion by osteoblasts. IL-6 is a multifaceted cytokine affecting pro- and anti-inflammatory signaling26. It was demonstrated that IL-6 is crucial for bone loss in estrogen deficiency and inflammatory bone loss in rheumatoid arthritis27,28. Furthermore, serum IL-6 correlated with bone turnover rate in MM patients29. We treated osteoblasts with PROS1 and detected as expected decreased protein levels of active β-Catenin indicating decreased osteoblast differentiation induced by PROS1-MERTK-RHOA axis as previously shown16,30 (Fig. 4H). Concomitantly, PROS1 led to inhibition of pro-osteoblastic downstream signaling pathways ERK and AKT (Fig. 4H). It was shown that SOCS1 negatively regulates IL-6 production31. Therefore, we hypothesized that PROS1 might affect SOCS1 expression in osteoblasts. Interestingly, we detected decreased Socs1 and Socs3 expression in osteoblasts upon stimulation with PROS1, suggesting that PROS1 downregulates SOCS proteins in osteoblasts leading to pro-inflammatory IL-6 cytokine signaling in osteoblasts (Fig. 4I and J). Subsequently we found increased STAT1 signaling, which likewise increases IL-6 production32 (Fig. 4H).
PROS1 inhibits IL-6 signaling in osteoclast precursors by upregulation of SOCS expression to induce osteoclast differentiation
It is widely assumed that IL-6 increases bone turnover and osteoclast formation indirectly via upregulation of RANKL expression in osteoblasts and osteocytes, because it was shown that IL-6 exerts direct inhibitory effects on osteoclast differentiation in vitro and in vivo33,34,35,36. As TAM receptors exert potent functions in macrophages and are linked with osteoclastic bone resorption in vitro, we sought to investigate the interplay of PROS1 and IL-6 also in osteoclastogenesis17,37. We performed osteoclast differentiation assays using BM macrophages from C57BL/6J mice. We observed that IL-6 blocks osteoclast differentiation in baseline. Albeit, in presence of 100 nM PROS1, IL-6 only slightly decreased osteoclast numbers (Fig. 4K and L), indicating that PROS1 potently blocks IL-6 - mediated inhibition of osteoclast differentiation of BM macrophages in vitro. It was shown that SOCS1 and SOCS3 regulate osteoclast differentiation by blocking inhibitory effects of inflammatory cytokine signaling by IFN and IL-6 on RANKL-mediated osteoclast differentiation signals at inflammation sites36. Additionally, IL-6 strongly suppressed TRAF6 and IκB expression levels and RANKL-induced IκB phosphorylation in osteoclasts in absence of SOCS336. We sought to elucidate if PROS1 inhibits IL-6/IL-6R/gp130 downstream pathways via upregulation of SOCS proteins. Indeed, we detected increased Socs1/3 expression in osteoclast precursor cells upon stimulation with PROS1 (Fig. 4M and N). The cell differentiation fate of myeloid precursor cells into macrophage subtypes or osteoclasts relies on the cytokine milieu in the BM, particularly in diseases with pathological osteoclast formation38. Our data indicates that PROS1 inhibits proinflammatory, anti-osteoclastogenic IL-6 signaling in myeloid precursor cells and macrophages, thereby inducing a shift towards osteoclast differentiation in these cells. In line with this, immunofluorescence staining of osteoclast cultures showed that IL-6 induced typical macrophage morphology despite presence of M-CSF and RANKL and this process could be inhibited by PROS1, leading to osteoclast, but not macrophage formation (Fig. 4O, arrows).
PROS1 fuels myeloma bone disease
Based on our results, we propose that PROS1 might be a driver of MM bone disease by suppressing osteoblast function and promoting osteoclast differentiation and osteolysis, by inhibiting IL-6 mediated pro-inflammatory cytokine signaling in BM macrophages or myeloid osteoclast precursors by induction of SOCS proteins. To investigate this hypothesis, we co-cultured myeloma cells overexpressing PROS1 with BM macrophages. Intriguingly, we observed increased myeloma - induced osteoclast differentiation mediated by PROS1 overexpression(Fig. 5A and B).
To study the role of PROS1 in myeloma bone disease in vivo, we injected JJN3 MM cell line with enforced expression of PROS1 into NSG mice (SFig. 1B). We used the TAM receptor - independent cell line JJN3 to minimize detrimental effects of PROS1 on MM plasma cells. μCT 3D reconstructions of the tibiae showed increased bone destruction in the metaphysis of mice injected with JJN3 PROS1+ (Fig. 5C). Quantification analysis revealed decreased bone volume induced by reduced trabecular number, indicating increased osteolysis (Fig. 5D-G). MM infiltration measured by FACS did not show significant differences in MM tumor load (Fig. 5H).
Histomorphometric analysis revealed increased osteoclast numbers and decreased osteoblast numbers in mice injected with PROS1 overexpressing JJN3 cells (Fig. 5I-K). The osteoclast marker Tartrate-resistant acid phosphatase (TRAP) 5b was increased in serum of mice injected with PROS1+ JJN3 cells (Fig. 5L). In contrast, bone formation marker Procollagen I amino-terminal propeptide (P1NP) was not increased in comparison to mice injected with control-transduced JJN3 cells (Fig. 5M). All in all, we identified PROS1 as a driver of MM bone disease by promoting osteoclast differentiation and bone destruction in vivo. We could confirm these results using the MERTK small molecule inhibitor R99216. R992 did not alter JJN3 MM cell infiltration in the BM, confirming that MERTK is redundant for JJN3 tumor progression in vivo (Fig. 6A). Albeit, we detected reduced myeloma bone disease by μCT analysis in tibia of R992 treated mice (Fig. 6B-F). Importantly, R992 led to prolonged overall survival (Fig. 6G).
{kind=link}