Phylogenomic characterisation, diversity, and distribution of UBA12189
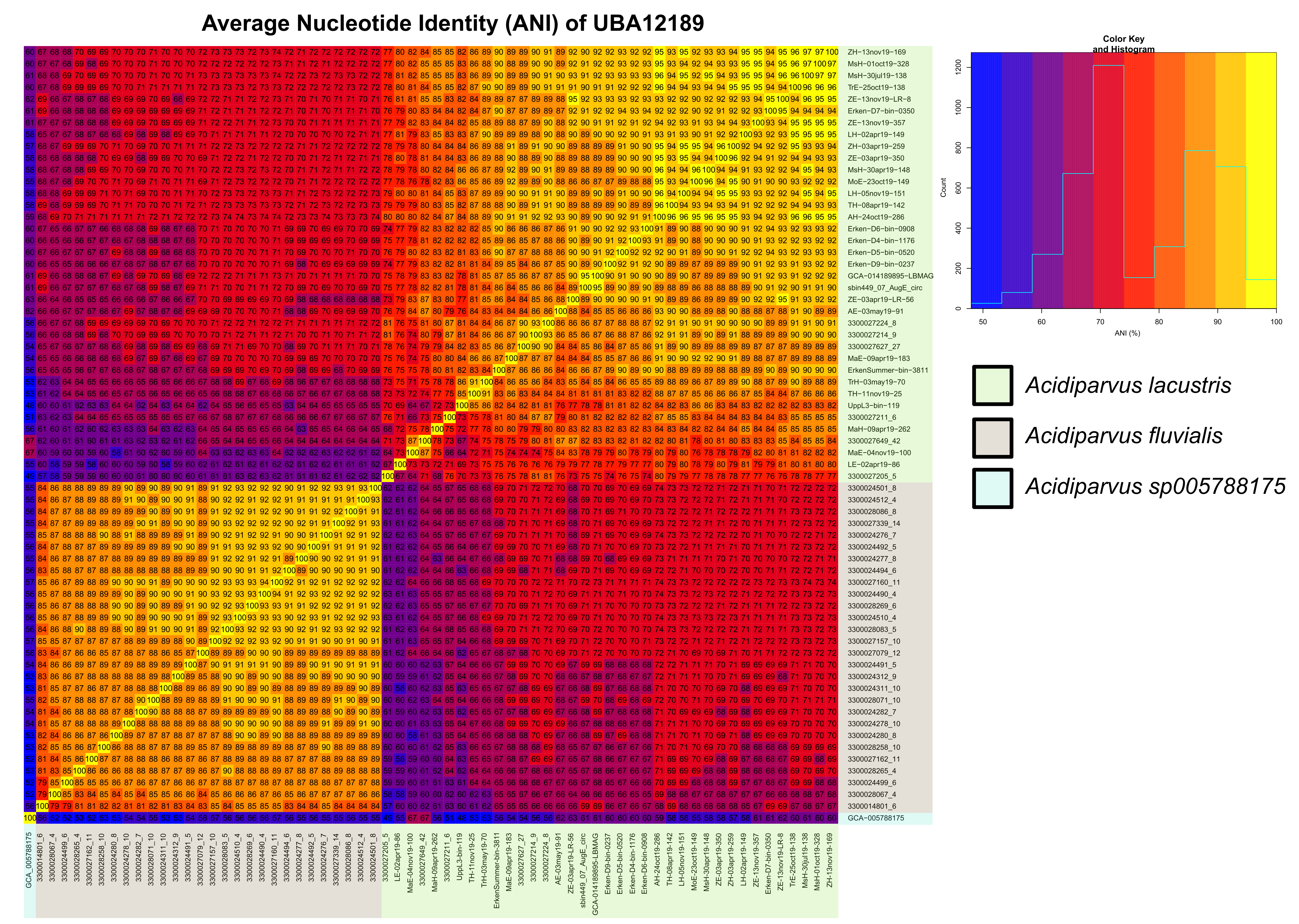
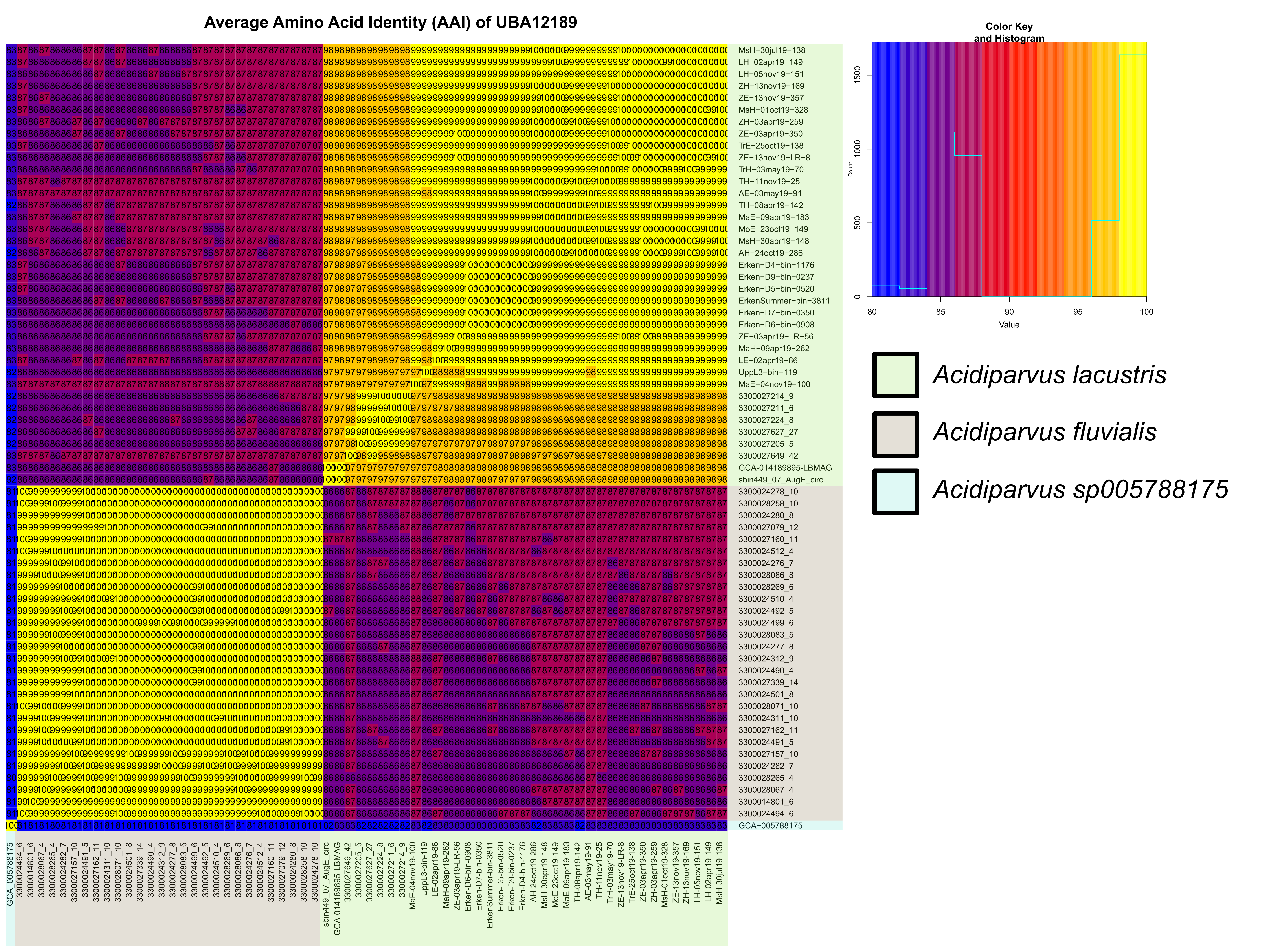
A total of 66 MAGs (63 MAGs from short-read sequencing and 3 MAGs from long-read sequencing) of the genus UBA12189 were investigated in this study. This genus, alongside Holophaga, Geothrix and four novel genera (JADJVL01, JAFDVN01, JACQZU01 and WRHW01) comprise the family Holophagaceae. All MAGs have > 70% completeness and < 5% contamination, including two complete, circular genomes (ZE-13nov19-LR-8 and sbin449) (Supplementary Table 5). Most of the MAGs were obtained from freshwater environments, except for 4 MAGs assembled from the Columbia River estuary. All the MAGs have streamlined genomes, with small, estimated genome sizes ranging from 1.12 Mb to 1.37 Mb, and low GC contents ranging from 44.3–48.7% (Supplementary Table 5). These are the smallest recorded genome sizes of Acidobacteriota so far and this lineage is the only genus with streamlined genomes under the family Holophagaceae. A phylogenetic tree of all available Holophagaceae genomes was constructed with Hydrogenedentota as the outgroup, where three separated clades of UBA12189 can be observed (Fig. 1, Supplementary Table 5). Interestingly, MAGs isolated from the current study are in the same phylogenetic clade (species I) together with MAGs isolated from Lake Biwa and estuarine environment (Fig. 1), while species II contains MAGs exclusively from rivers and species III consists of one MAG assembled from alpine Lake Fuxian. Average nucleotide identity (ANI) and average amino acid identity (AAI) also suggest the same results, showing three different species (ANI < 95%) nonetheless the same genus (AAI > 65%) [73] (Supplementary Fig. 2, Supplementary Fig. 3). We therefore propose that UBA12189 is a novel genus consisting of three species that we tentatively name ‘Ca. Acidiparvus’. Species I and II are designated ‘Ca. A. lacustris’ and ‘Ca. A fluvialis’, because of their occurrence in freshwater lakes and rivers, respectively. Species III did not meet the criteria to be named as Candidatus species; we thereby refer to it as Acidiparuvs sp005788175 based on current GTDB classification (Supplementary Information).
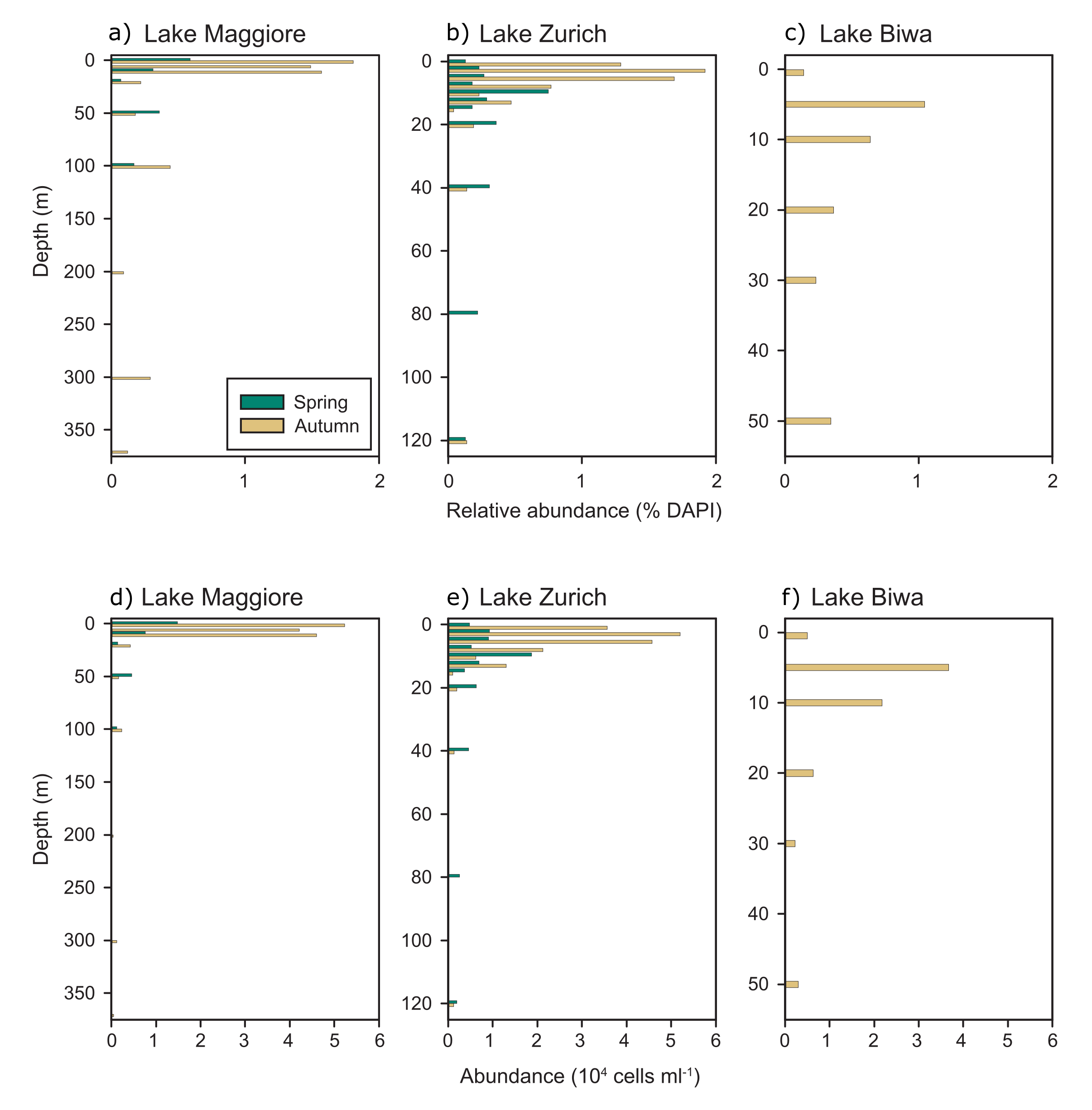
To estimate the abundance of Acidiparvus in freshwater metagenomes, metagenomic fragment recruitment was employed. Coverage per Gb ranges from 0 to 13 (Supplementary Table 2), with the highest coverage obtained from the hypolimnion of Lake Biwa during mixis in Spring (March-April) and the epilimnion in Autumn and Winter (October and February). On average, Acidiparvus is most abundant in Lake Biwa (Japan), followed by Lake Maggiore (Italy) and Lake Zurich (Switzerland). The results suggest that Acidiparvus is a flexible genus that can inhabit both the surface waters and hypolimnion water columns of large lakes across distant geographical locations. Furthermore, results show that only A. lacustris is abundant in the lakes sampled in the current study, while A. fluvialis and A. sp005788175 are practically absent, probably due to A. fluvialis being found exclusively in rivers (Fig. 1). Genome replication rates in freshwater UBA12189 were measured with GRiD based on ori/ter values, ranging from 1.1 to 1.56, indicating slow growth and replication rates at the sampling timepoints (Fig. 1, Supplementary Table 3).
Acidiparvus appears to be free-living
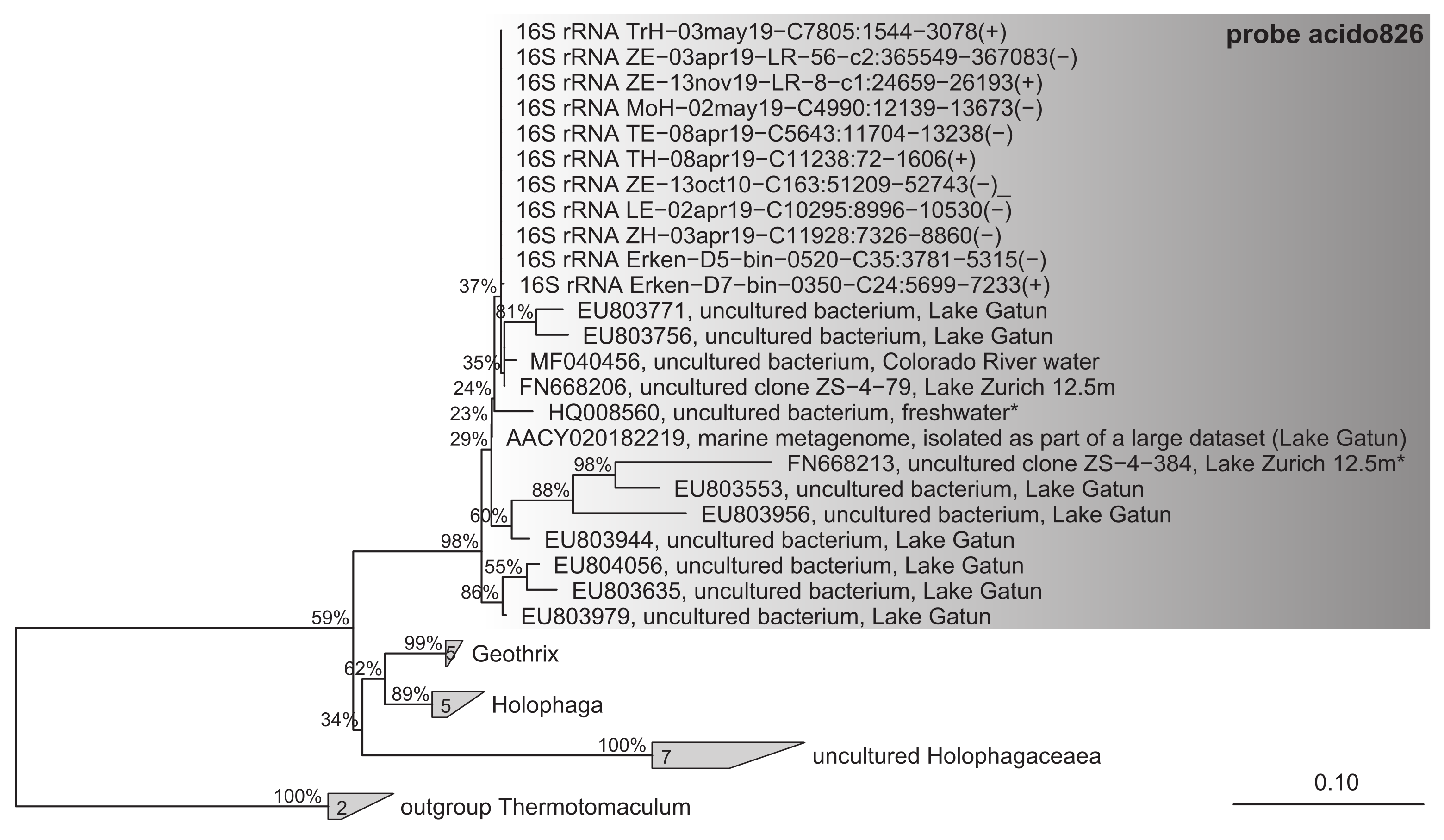
CARD-FISH with probe acido826 show that Acidiparvus can account for up to 1.9% of all prokaryotes with abundances of up to 5.2x104 cells ml− 1 in lakes Zurich, Maggiore, and Biwa (Fig. 2, Supplementary Fig. 4). Acidiparvus have maximum abundances in the epilimnion and gradually decreases with depth with higher densities during autumn compared to spring. CARD-FISH microscopy images show that all Acidiparvus are very small and free-living, they were never found attached or in close vicinity to other microorganisms (Fig. 3, Supplementary Fig. 5). To further examine if they have close association with other microorganisms, we performed BLASTP to identify any traces of horizontal gene transfer (HGT). All but one MAG (UppL3-bin-119) have > 90% of the genome of Acidobacteriota origin (to be specific, UBA12189), suggesting no evidence of potential recent HGT events (Supplementary Table 6). Furthermore, without a complete type II secretion system (T2SS), and the complete absence of genes involved in type III, type IV and type VI secretion systems, there is no concrete evidence of any potential host association (Supplementary Information). This reinforces the free-living aspect of Acidiparvus. However, we found extremely low metabolic versatility in the MAGs suggesting this bacterium cannot live alone, which will be described in detail in the following paragraphs.
The genomic repertoire of Acidiparvus is characterized by low metabolic versatility and a high extent of auxotrophy
We conducted comparative genomic analyses of 66 Acidiparvus genomes, including two circular, complete genomes (ZE-13nov19-LR-8 and sbin449), to obtain a comprehensive view on the genomic repertoire and metabolic potential of this Acidobacteriota lineage (Fig. 4, Fig. 5). Since Acidiparvus have extremely limited metabolic capacity, we compared their genomes to other aquatic genome streamlined microbes such as Nanopelagicales [10] and Methylopumilus [74] in this study.
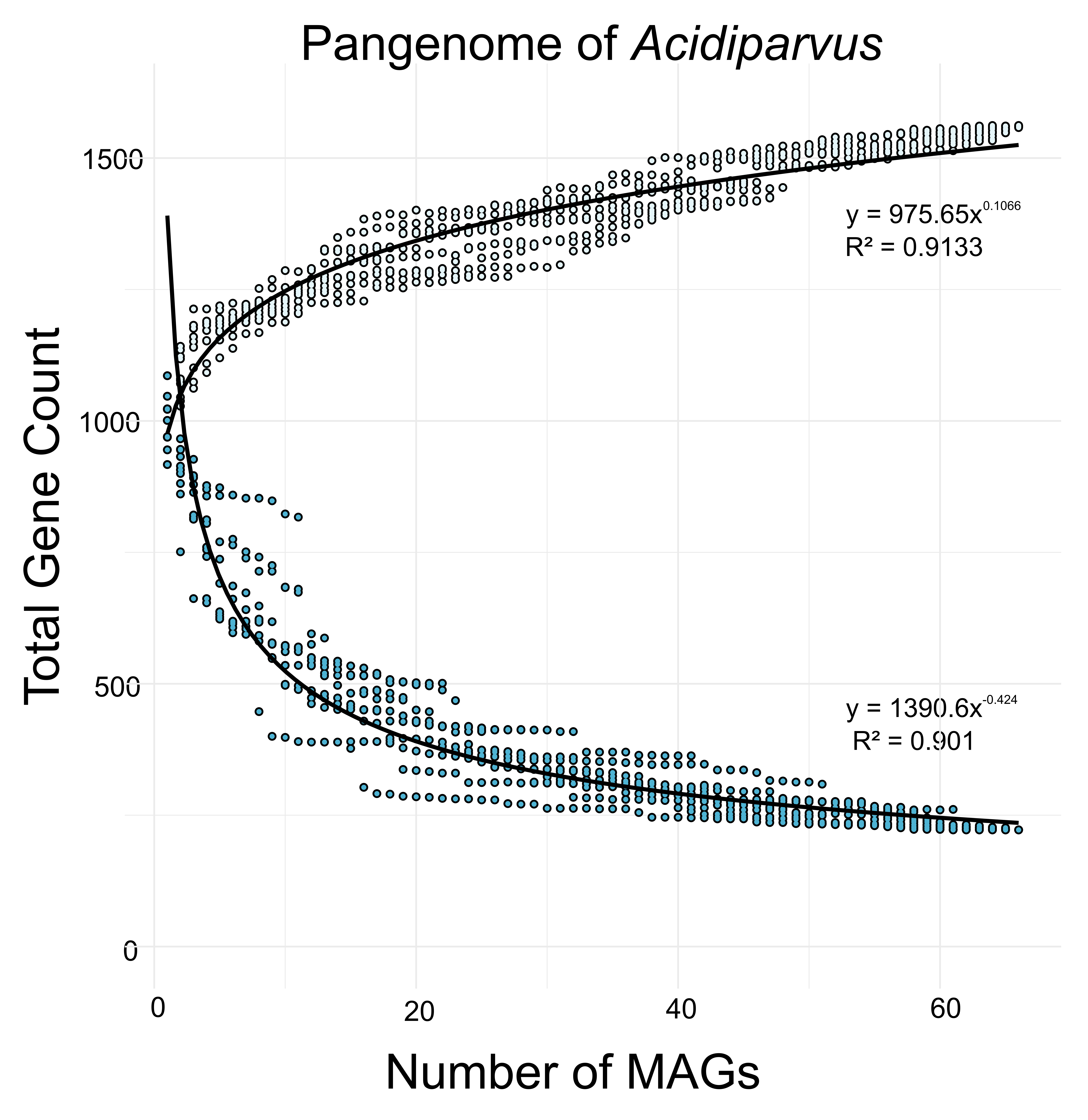
All genomes of Acidiparvus contain a relatively high fraction of hypothetical proteins comprising 10% − 14% of the genomes (Supplementary Table 5), which is comparably higher than Nanopelagicales (6% − 10%) and Methylopumilus (7% − 10%). The core-genome of the 66 Acidiparvus MAGs encodes only 222 genes (18%-28% of the assembled genomes), suggesting a higher proportion of flexible genomes with highly diverse auxiliary genes. The pangenome is estimated to contain 1562 genes and the power law regression analysis suggested an almost saturated pangenome for this genus (Supplementary Fig. 6, Supplementary Table 7). Core genes in Acidiparvus only comprise 14% of the predicted pangenome, which is significantly lower than that of Nanopelagicales and Methylopumilus (29% and 48% respectively), suggesting a high genetic diversity for adaptation during genome streamlining [10, 74]. Within the family Holophagaceae, Acidiparvus shares only 12 core genes with the four other undescribed genera (JACQZU01, JADJVL01, JAFDVN01 and WRHW01) and 6 core genes with Holophaga and Geothrix, indicating that Acidiparvus is highly divergent and metabolically distant from other Holophagaceae lineages.
Limited capacity for amino acid and vitamin biosynthesis and membrane transporters
Acidiparvus have highly restricted capacities for amino acid biosynthesis, which are limited to cysteine, arginine and proline. When compared to the rest of Holophagaceae and other genome streamlined freshwater taxa, Acidiparvus is auxotrophic for the majority of amino acids. Sixty-three MAGs contain genes for proline synthesis while 54 MAGs are able to produce cysteine (Fig. 6). Over half of the MAGs have the complete pathway for arginine synthesis while the others are lacking amino-acid N-acetyltransferase to convert glutamate to N-acetylglutamate. Compared to Nanopelagicales and Methylopumilus, Acidiparvus is the only genome streamlined genus to encode N-acetylornithine carbamoyltransferase (argF) and acetylornithine deacetylase (argE), allowing them to synthesize arginine from carbamoyl-P.
Cysteine is synthesized by cystathionine gamma-lyase instead of cysteine synthase, thus no hydrogen sulfide is incorporated into the cells. Since no evidence of genes responsible for sulfate reduction or assimilation were identified, the mode of acquisition of reduced sulfur is unknown. As a notable difference from streamlined Nanopelagicales and Methylopumilus, Acidiparvus may take taurine and sulfonate as an external sulfur source since half of the MAGs encode sulfonate/Nitrate/Taurine transporters. However, taurine/sulfonate degradation genes were not identified.
Most of the MAGs share transporters for phosphate, molybdate, sodium, phospholipid and cobalamin while afu iron(III) transporters are exclusively found in A. sp005788175 (Fig. 6, Supplementary Table 8). Half of the MAGs have sulfonate/Nitrate/Taurine transporters albeit lacking substrate-binding protein (ssuA), indicating taurine and sulfonate as potential sulfur sources. However, as mentioned above, taurine/sulfonate degradation genes are absent in the MAGs. Compared to Nanopelagicales and Methylopumilus, Acidiparvus is the only genome streamlined lineage to encode sodium and molybdate transporter systems (natAB, modABC) (Fig. 6).
We screened the MAGs for high-affinity pstABCS transport systems for phosphorus acquisition. We found 57 MAGs encoding the complete set of pst transport system, while it was lacking in 4 MAGs (Fig. 6, Supplementary Table 8). This suggest that this lineage is well adapted since phosphorus is the primarily limiting nutrient in freshwater lakes [76, 77]. All Acidiparvus MAGs encode for cobalamin (vitamin B12) transporter (btuB), with 53 MAGs having 2 copies of the gene. However, the substrate-binding protein for cobalamin (btuF) is exclusively found in A. lacustris MAGs. Major facilitator superfamily transporter (MFS) for nitrate/nitrite was only found in 3 MAGs, suggesting alternative sources of nitrogen uptake. A few MAGs encode for MFS bidirectional aromatic amino acid transporters (4-hydroxybenzoate, benzoate, cis, cis-muconate, 3-hydroxyphenylpropionic acid). However, genes associated with benzoate degradation are absent in these MAGs, raising the question of the aim of importing aromatic compounds. Phenylacetic acid degradation protein (paaY) was identified in all MAGs, suggesting the bacteria’s ability to break down aromatic compounds.
Given that all Acidiparvus lack the ability to produce most amino acids, they must rely on external sources. However, amino acid transporters from the ABC family (livGFHMK, oppABCDF, dppABCDF) are absent in all but one MAG, namely the complete genome from lake Biwa (sbin449). Nonetheless, the substrate-binding protein (livK) is missing in this MAG. In comparison, other genome streamlined bacteria such as Nanopelagicales contain branched amino-acid transporters (livGFHMK), while these transporters are absent in Methylopumilus but they harbour complete amino acid biosynthesis gene sets (Fig. 6) [74]. Despite Acidiparvus lacking amino acid transporters, they encode homologues of oligopeptide transporters of the OPT superfamily (COG1297) and proton-dependent oligopeptide transporters of the POT superfamily (TC.POT; K03305) (Fig. 6). These putative oligopeptide transporters are also identified in all other Holophagaceae lineages but not in Nanopelagicales and Methylopumilus. Different from other prokaryotic OPTs which usually have 16 transmembrane domains, most of the OPT homologues in Acidiparvus harbour 17 transmembrane domains (Supplementary Table 10). It was suggested that this transporter can act as both peptide and iron-siderophore transporters [78]. Most of the Acidiparvus MAGs also encode for multiple copies of TC.APA basic amino acid/polyamine antiporters (K03294), allowing them to export polyamines in exchange of amino acids. Indeed, all Acidiparvus MAGs have at least one set of genes for polyamine production (speAB, agmatine and putrescine production), enabling them to potentially produce and exchange polyamine for amino acids (Fig. 6). As for nitrogen uptake, urea transporter and cyanophycinase are absent in all Acidiparvus genomes, indicating that these are not the major nitrogen sources. Most of the MAGs encode glutamate dehydrogenase (gdhA), which allows them to convert ammonium to glutamate as a source for nitrogen assimilation. However, ammonium transporters (amt) were not identified in any of the Acidiparvus genomes, suggesting the presence of putative novel, currently unannotated proteins to import ammonium across the cell membrane.
As described above, Acidiparvus have limited number of cell membrane transporters, averaging only 48.5 transporters per genome (46.1 transporters per Mb) (Supplementary Table 11). It appears that other members of the family Holophagaceae also have lower proportion of transporters, with Holophaga and Geothrix having only 57.9 and 38.9 transporters per Mb respectively. Yet, the total number of transporters per genome is much higher (215 and 127 transporters, respectively) due to their larger genome sizes and they encode a very versatile set of genes for various metabolisms. When compared to other genome streamlined lineages, Nanopelagicales and Methylopumilus have a higher number of transporters to compensate for auxotrophy (Supplementary Table 11). With the great extent of auxotrophy and higher proportion of hypothetical proteins, it is likely that Acidiparvus have novel, unannotated transporters for nutrient acquisition.
Apart from being auxotroph for most of the amino acids, Acidiparvus are also auxotrophs for vitamins B1 (thiamin), B3 (nicotinamide adenine dinucleotide), B6 (pyridoxine), B12 (cobalamin), and B7 (biotin). However, Acidiparvus have complete biosynthesis pathways for vitamin B2 (riboflavin) and vitamin K2 (menaquinone) (Fig. 6). Not only does Acidiparvus have limited metabolic capacities and high degree of auxotrophy as described above, this Acidobacteriota lineage also lacks hydrogenases, cyanophycinases, CRISPR-Cas systems and biosynthetic gene clusters (BGC). With a very limited genome repertoire, as well as the small number of cell membrane transporters, this all points to a scenario where free-living Acidiparvus have a constrained cell metabolism that is highly specialized [6]. With no specific co-occurring microbes identified, Acidiparvus is suggested to be a free-living scavenger in the lakes. Although canonical genes for various metabolic pathways cannot be identified, the high proportion of unannotated genes and auxiliary genes in the pangenome may offer the flexibility for Acidiparvus to survive.
Unique set of genes in Acidiparvus as adaptation to the sunlit epilimnion of lakes
Since Acidiparvus has low metabolic versatility and appears to lack several general molecular machineries for survival, we compared the genomes with other genera within the family Holophagaceae (Geothrix, Holophaga, JAFDVN01, JADJVL01, JACQZU01 and WRHW01) and other genome streamlined freshwater microbes to identify unique features of this genome streamlined genus.
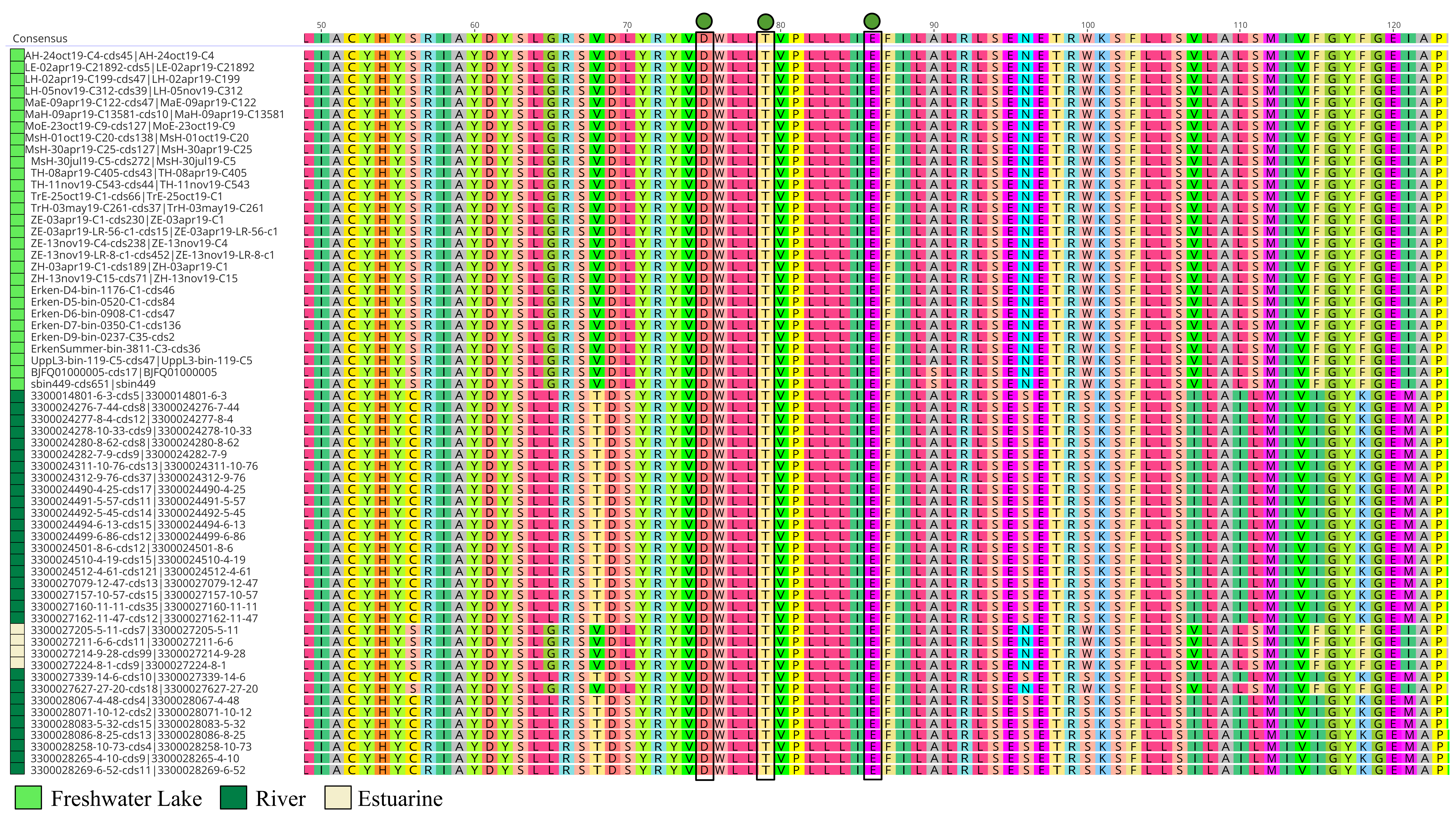
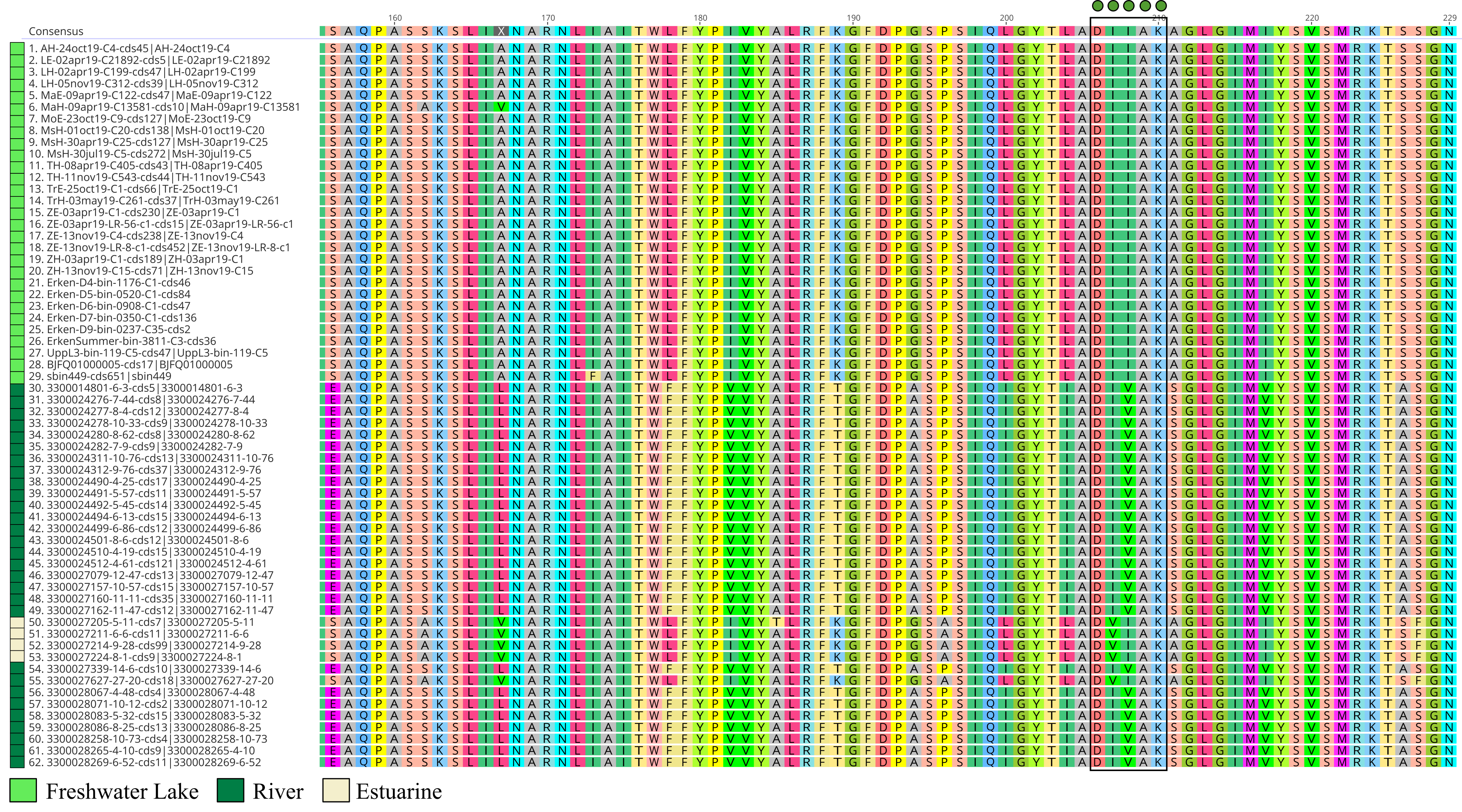
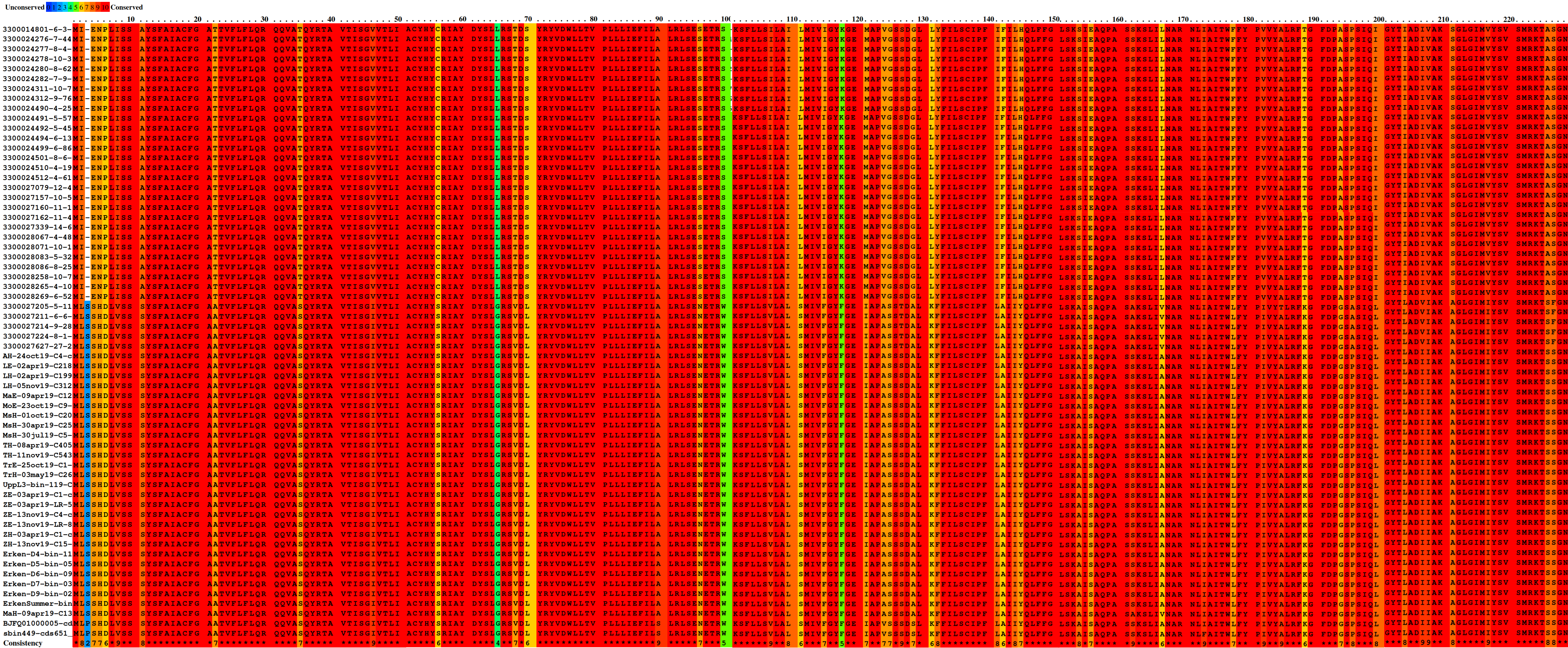
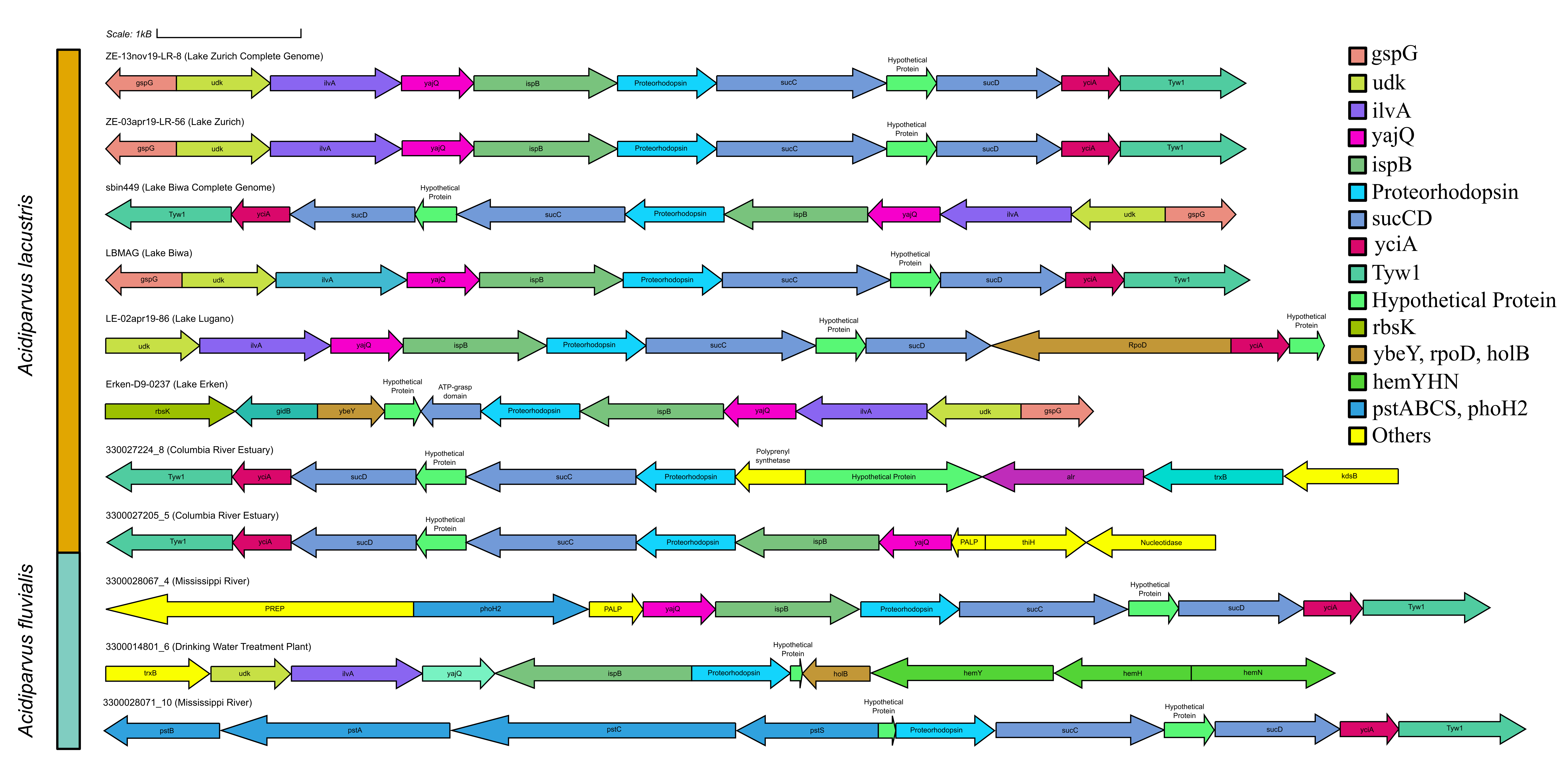
Microbial rhodopsins are transmembrane proteins that are light driven proton pumps for specific biological functions such as ATP generation, which is an adaptational feature in genome streamlined bacteria found in the pelagial of oceans and lakes [79, 80]. Upon screening for rhodopsins in all Holophagaceae MAGs, Acidiparvus is the only genus in this family to encode for rhodopsins. A total of 62 rhodopsins were identified in the 66 MAGs. All the rhodopsins were predicted to be proteorhodopsins harbouring DTE-motifs in helix C, which appear to form their own phylogenetic cluster branching off proteorhodopsins from other prokaryotic taxa, while phylogenetically closest to Alpha- and Gammaproteobacterial DTE-motif proteorhodopsins (Fig. 7). All proteorhodopsins have L (leucine) at amino acid position 105 and retinal-binding lysine in helix G, indicating they are green-light absorbing and light sensitive (Supplementary Fig. 7, Supplementary Fig. 8) [81]. Despite the MAGs were sourced from different geographical locations, the rhodopsins are highly conserved (Supplementary Fig. 9) and flanked by highly conserved Acidiparvus-like genes (Supplementary Fig. 10, Supplementary Table 12, Supplementary Table 13). This suggests that the rhodopsins were not acquired through recent horizontal gene transfer events. As the only lineage under this family with streamlined genomes and reduced metabolic capacity, proteorhodopsins may play a role in their survival through augmenting a proton gradient and likely ATP synthesis [79]. A photoheterotrophic lifestyle is commonly reported for other genome streamlined freshwater lineages such as Nanopelagicales, Methylopumilus and Fonsibacter (maximum genome size of 1.4 Mbp, 1.35 Mbp and 1.16 Mbp respectively) [3, 6, 10, 74]. However, Nanopelagicales, Methylopumilus and Fonsibacter all harbour genes necessary for retinal biosynthesis (crtY, crtI, crtB, crtE), while these genes are absent in all Acidiparvus MAGs. Despite the lack of retinal chromophore, recent studies showed that rhodopsin can still function through the uptake of retinal from the environment [82] or by a putative unknown retinal biosynthesis pathway [83].
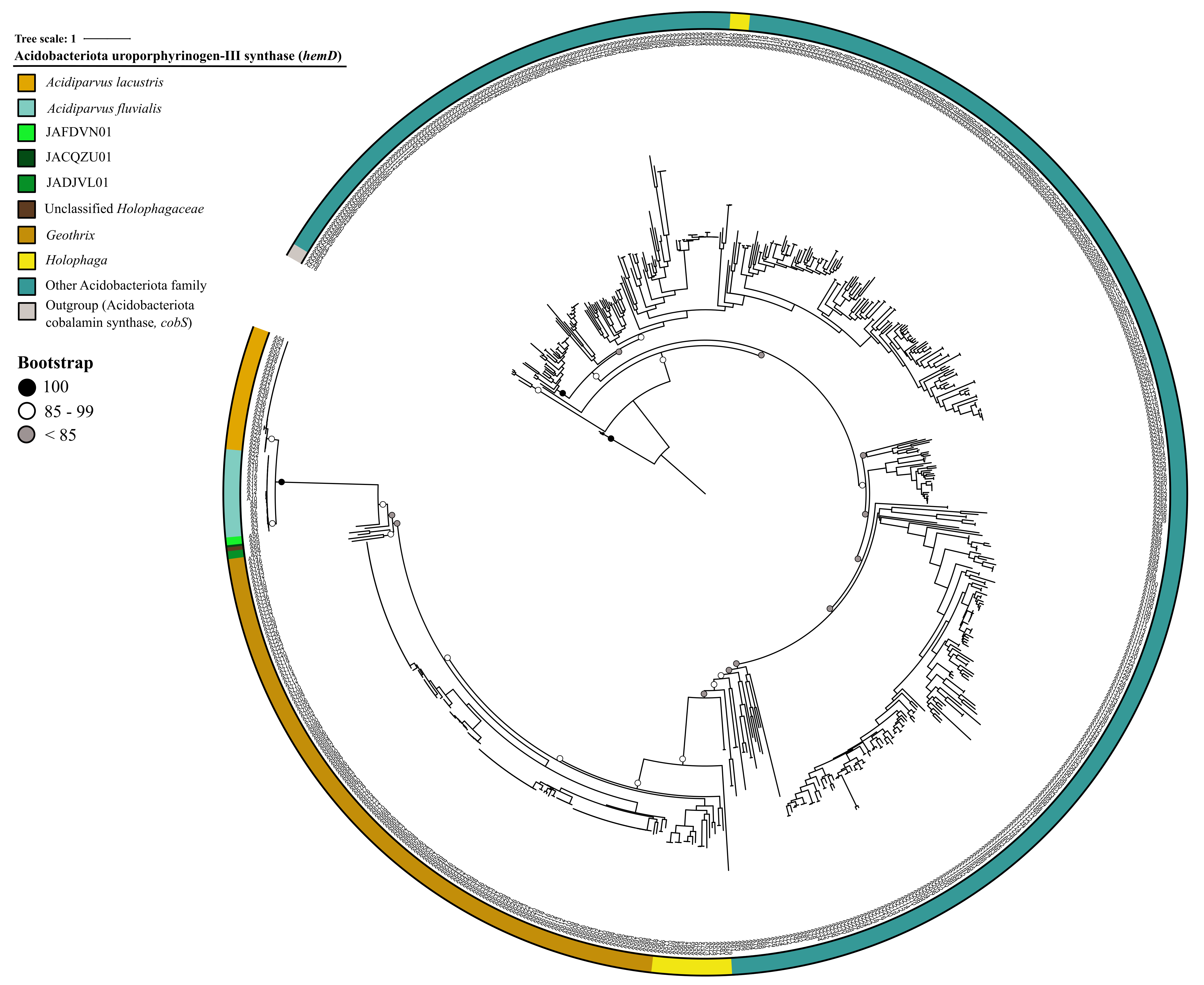
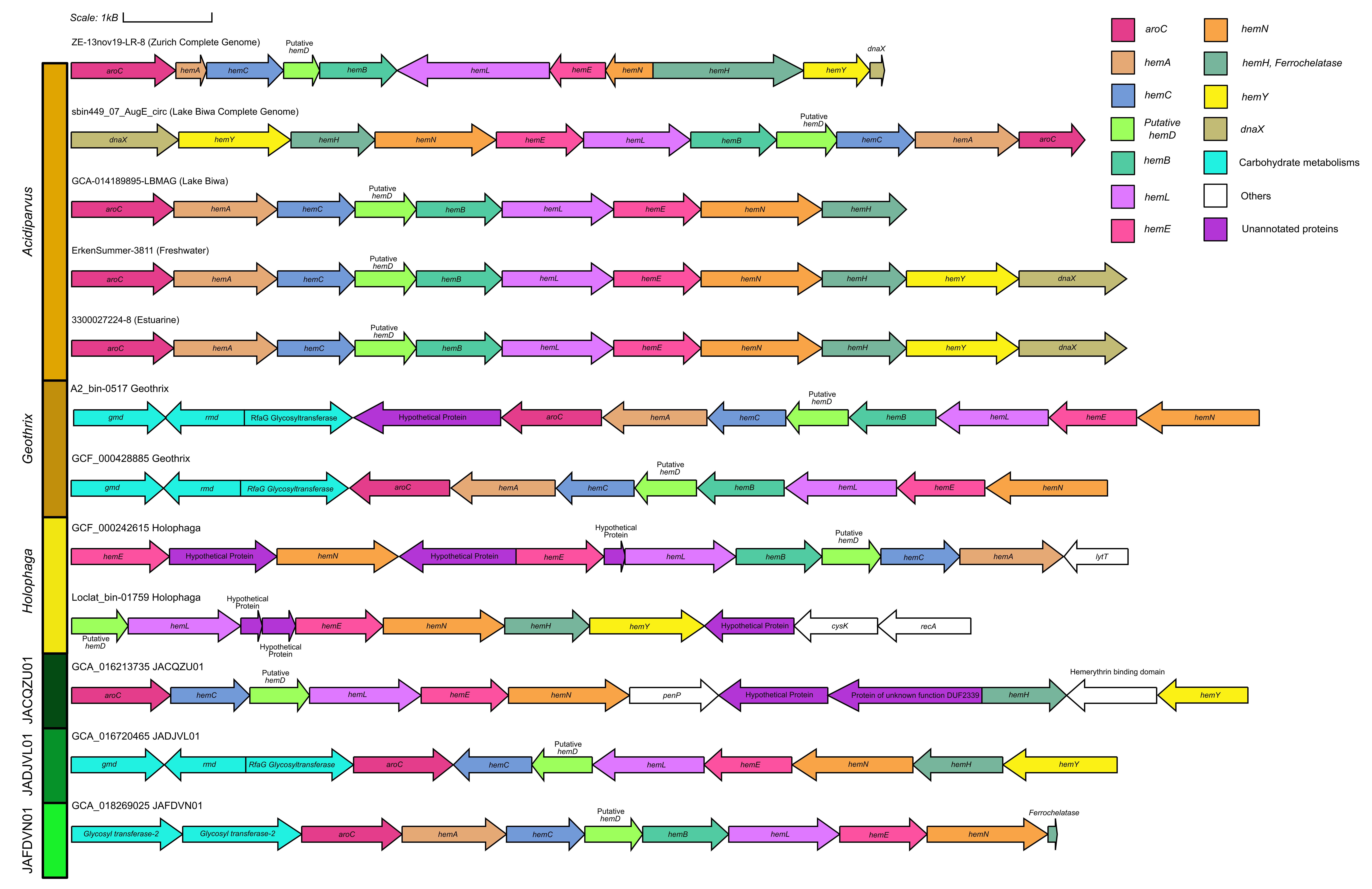
Surprisingly, despite having extremely limited metabolic versatility, a complete protoporphyrin-dependent (PPD) heme biosynthesis pathway was found in most of the MAGs in A. lacustris and A. fluvialis (Fig. 6). This might suggest that Acidiparvus may have symbiotic relationships with other microorganisms exchanging heme for nutrients, though it appears to be free-living from CARD-FISH microscopy images (Fig. 3, Supplementary Fig. 5). A recent study showed that heme auxotrophy is widespread across abundant microbial lineages in aquatic environments, indicating heme reliance in these ecosystems and suggesting heme as public goods in microbial interactions [84, 85]. However, subunits of heme exporters (ccmABCD) are only identified in three MAGs, suggesting biosynthesized heme is not exported, or this genus encodes novel, unannotated heme exporters. Although heme biosynthesis is common to most Holophagaceae, Acidiparvus has hemD (uroporphyrinogen-III synthase) genes that are phylogenetically closer to the genera JADJVL01, JACQZU01 and JAFDVN01, forming a monophyletic cluster, indicating hemD is well conserved throughout the lineage (Supplementary Fig. 11). Similar to proteorhodopsins, heme biosynthesis gene clusters are highly conserved (Supplementary Fig. 12) and not flanked by genes from other taxa (Supplementary Table 14), indicating that heme biosynthesis is not a result of recent horizontal gene transfer. It is hypothesised that heme production among Acidiparvus occupies a certain ecological niche in freshwater systems since the whole pathway was kept intact during genome streamlining. Although no heme exporters were identified, heme may be released upon phage lysis. Most of the MAGs also encode for heme-requiring enzymes such as cbb3-type cytochrome c oxidase, catalase, peroxidase and succinate dehydrogenase [85], suggesting these genes give Acidiparvus a selective advantage during genome streamlining given this bacterial taxon became auxotrophic to most amino acids and vitamins.
Furthermore, most of the MAGs (61 out of 66) encode both class I cyclopyrimidine dimer photolyase (phrB) and catalase-peroxidase (katG), both with the ability of using light energy to repair UV-induced DNA damage, suggesting that Acidiparvus is adapted to shallow depths in the water column. This is consistent with metagenomic fragment recruitment and CARD-FISH counts as they are abundant in the epilimnion of lakes, while also found in the deeper hypolimnion during mixis. These genes also enable Acidiparvus to avoid heme toxicity since most MAGs have complete de novo heme biosynthesis pathway, which can produce reactive oxygen species (ROS) as byproducts [85–87]. Moreover, the detoxification ability of these genes may function as public goods in microbial interactions [84].
Sixty-three MAGs encode subunits for all five complexes of the electron transport chain (ETC), with cbb3-type as complex III. Interestingly, 63 out of 66 MAGs harbour cbb3-type cytochrome c oxidase subunits I-III (ccoN, ccoO, ccoP), forming the minimal functional unit [87] (Fig. 6, Supplementary Table 8). These genes were not found in other Holophagaceae and any other genome streamlined lineages such as Nanopelagicales, Methylopumilus and Patescibacteria [5, 10, 89]. These oxidases have high oxygen affinity, indicating a putative capacity for microaerobic respiration in low O2 environments, conserving energy under such conditions [28, 90, 91]. These oxidases are suggested to have three roles for Acidiparvus. Firstly, conserving energy through O2 reduction in aerobic respiration. Secondly, adapting to microaerobic environments and O2 scavenging. Thirdly, expanding their distribution throughout epilimnion and hypolimnion columns [28, 89].
Besides the high-O2 affinity oxidases, 27 MAGs also encode for all subunits of the five complexes in the ETC, suggesting besides a fermentative lifestyle, Acidiparvus also participates in both O2 scavenging and aerobic energy production. Indeed, MAGs recovered from the hypolimnion from lakes Most, Thun and Maggiore (60m, 180m, 300m respectively) have both complete ETC and functional cbb3-type cytochrome c oxidase. Furthermore, CARD-FISH shows the presence of Acidiparvus down to a depth of 300m, and metagenomic fragment recruitment indicates that the lineage is abundant at both epi- and hypolimnion (Fig. 1, Supplementary Table 2). Thus, all evidence points towards a high flexibility of this lineage to adapt to both surface and deep water columns.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}