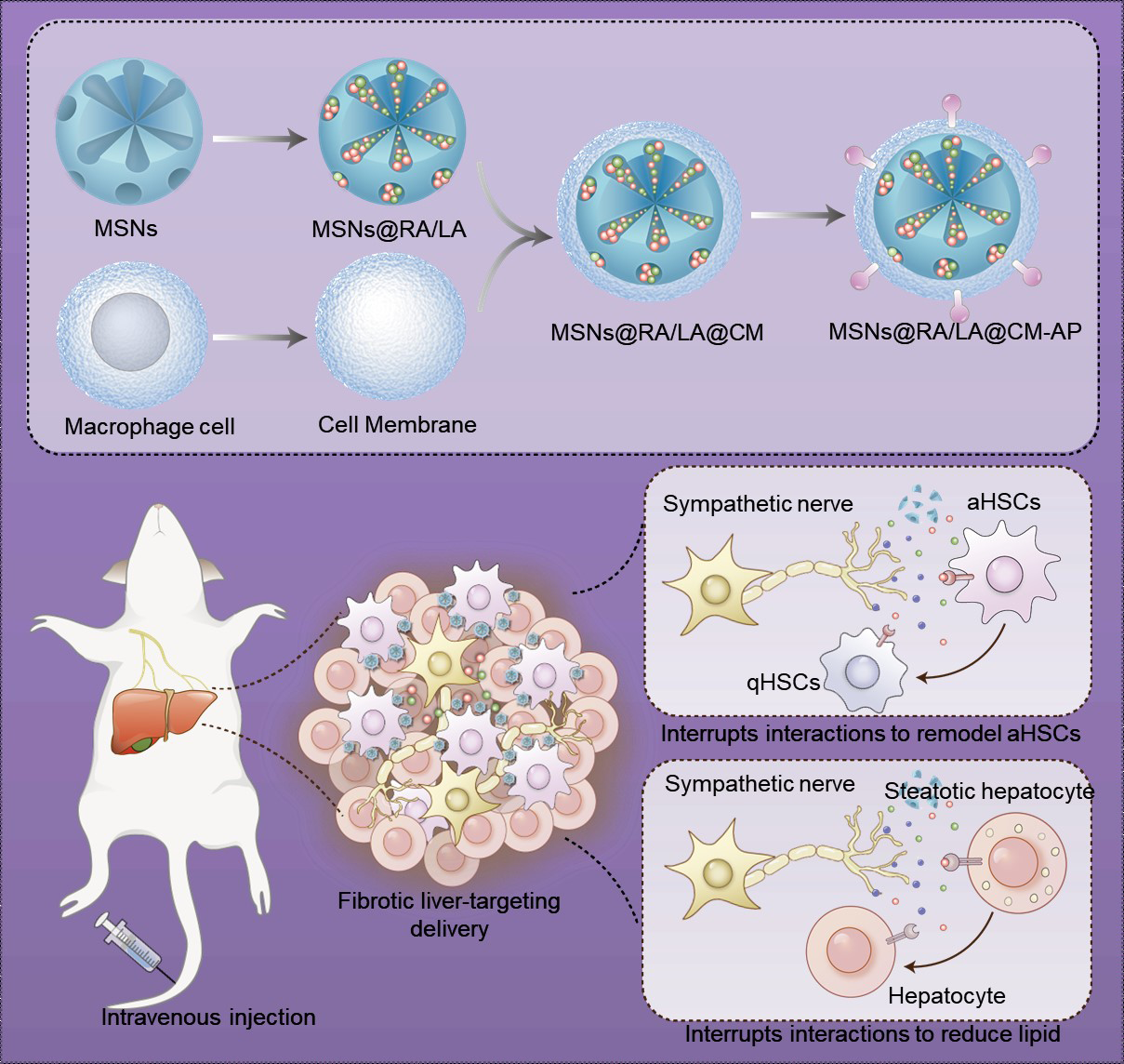
A previously reported sol-gel method was used to synthesize diselenide-bridged mesoporous silica nanoparticles (MSNs). Electron microscopy images (Fig. 1A) clearly revealed that the prepared MSNs had a uniformly spherical morphology with a size of approximately 80 nm. The N2 adsorption/desorption isotherms indicated a large surface area (487.2 m2/g), pore volume (0.41cm3/g), and average pore diameter (3.8) of the MSNs (Figure S1), which bestowed a high ability to preload RA and LA. The drug loading and cumulative release of RA/LA-loaded MSNs (MSNs@RA/LA) was measured by HPLC. The encapsulation efficiencies (EE%) of RA and LA was calculated to be 32.8 ± 2.0% and 25.1 ± 1.0%, while the drug loading content (DL%) of RA and Labetalol was 11.6 ± 0.64% and 9.1 ± 0.33%, respectively (Table S1). Considering that the fibrotic liver features excessive ROS, we investigated the ROS-responsive degradation and drug release of MSNs@RA/LA in a solution containing 100 × 10− 6 M H2O2. Due to the cleavage of the diselenide bridge, MSNs@RA/LA gradually collapsed into irregular aggregates in the presence of 100 × 10− 6 M H2O2 at 3 days (Fig. 1B and S2). Consistent with the degradation behavior, the cumulative release of RA and LA reached 61.1% and 66.5%, respectively, in H2O2-containing solution, whereas 7.9% RA and 11.2% LA were released from MSNs@RA/LA after 72 h in PBS solution. The ROS-responsive release was conducive to improving the drug release efficiency in fibrotic livers with high levels of ROS while minimizing drug side effects in normal tissues. Additionally, MSNs gradually consumed H2O2 as the concentration and time increased (Fig. 1C and S3), suggesting the ROS-scavenging ability of MSNs during the treatment of NAFLD.
To prolong the blood circulation time and decrease the capture of Kupffer cells, cell membranes were derived from macrophages and coated on the surface of RA/LA coencapsulated MSNs (MSNs@RA/LA to form MSNs@RA/LA@CM. Subsequently, CD44 aptamer-conjugated DSPE-PEG was directly fused with MSNs@RA/LA@CM using a noncovalent method. As shown in Fig. 1F, the formed biomimetic nanocomposites (MSNs@RA/LA@CM-AP) had a spherical structure with an MSN core enclosed by a thin membrane shell. DLS measurements revealed that MSNs@RA/LA@CM-AP had a slightly larger hydrodynamic size than did bare MSNs@RA/LA, and the surface potential decreased from − 6.2 mV for MSNs@RA/LA to − 21.3 mV due to the coating of negatively charged MSC membranes (Fig. 1G and H). These results indicated the successful coating of the cell membrane onto the nanoparticles. Additionally, the characteristic absorption of the CD44 aptamer at 260 nm was detected in the UV‒vis spectrum of MSNs@CM-AP, confirming the successful modification of the CD44 aptamer (Figure S4). Furthermore, MSNs@RA/LA@CM and MSNs@RA/LA@CM-AP displayed relatively good stability after 7 days of stockage in the cell medium, whereas aggregations were found on MSNs@RA/LA without cell membrane coating (Figure S5).
To explore their ability to target aHSCs, we investigated the cellular uptake of Cy3-labeled MSNs@RA/LA@CM-AP by TGF-β1-stimulated LX-2 cells via a laser confocal assay and flow cytometry. After treatment with TGF-β1 (2 ng/mL) for 24 h, the expression of fibrogenic markers such as α-SMA, Col1a1, Col3a1 and Col5a1 was significantly elevated (Figure S6), demonstrating the successful induction of aHSCs. Furthermore, our data showed the internalization of MSNs@RA/LA@CM-AP in aHSCs was remarkably greater than that of MSNs@RA/LA@CM and MSNs@RA/LA (Fig. 2A and C). However, the uptake of MSNs@RA/LA@CM-AP was low in quiescent HSCs (qHSCs), which was comparable to that of MSNs@RA/LA@CM (Fig. 2B and D). These results indicated the aHSC-specific targeting property of MSNs@RA/LA@CM-AP. To investigate the ability of MSNs@RA/LA@CM-AP to escape macrophages, MSNs@RA/LA, MSNs@RA/LA@CM and MSNs@RA/LA@CM-AP were incubated with RAW264.7 macrophages for 6 h. As shown in Figure S7, the uptake of MSNs@RA/LA@CM-AP and MSNs@RA/LA@CM was lower than that of MSNs@RA/LA in RAW264.7 cells, suggesting that the cell membrane coating might decrease the capture of Kupffer cells. Additionally, MSNs@RA/LA@CM-AP and MSNs@RA/LA@CM displayed a markedly increased blood retention than MSNs@RA/LA, which was likely due to the good colloidal stability and the immune-evasion propery of the macrophage membrane (Figure S8). Having demonstrated that the MSNs@RA/LA@CM-AP were able to target aHSCs in vitro, we investigated the tissue biodistribution of MSNs@RA/LA@CM-AP in liver-fibrosis mice induced by bile duct ligation (BDL) and in normal mice. We found that MSNs@RA/LA@CM-AP predominantly accumulated in liver tissues 8 h after intravenous administration (Fig. 2E and 2G). A quantitative assay indicated that MSNs@RA/LA@CM-AP showed greater accumulation efficiency in the fibrotic liver than in the normal liver, whereas MSNs@RA/LA@CM did not result in a significant difference in tissue accumulation between the fibrotic liver and normal liver, which could be explained by the greater uptake of MSNs@RA/LA@CM-AP by aHSCs in the fibrotic liver due to the modification of the CD44 aptamer. These results indicated the excellent ability of MSNs@RA/LA@CM-AP to actively target the fibrotic liver.
The biosafety of nanoparticles is an important concern for medical applications. We evaluated the cytotoxicity of these nanoparticles in LX-2 cells, HL7702 cells and HUVECs. Our data showed that the viability of these cells was above than 80% even when the concentration of MSNs and MSNs@RA/LA@CM-AP reached 100µg/mL (Figure S9A-C), indicating that MSNs@RA/LA@CM-AP had a low effect on cell survival. We subsequently evaluated the systemic toxicity of MSNs@RA/LA@CM-AP in the mice. Encouragingly, repeated administration of MSNs@RA/LA@CM-AP did not induce significant changes in body weight, blood biochemistry, or organ histopathology compared with those of the control group (Figure S9D-F). These results suggested that MSNs@RA/LA@CM-AP were a low-toxicity nanocarrier system.
To investigate the antifibrotic activity of MSNs@RA/LA@CM-AP, we detected the expression of fibrogenic markers in aHSCs after treatment with various formulations. QT-PCR revealed that the expression of α-SMA, Col1a1, Col3a1 and Col5a1 was obviously downregulated following treatment with free RA, free LA, RA plus LA MSNs@RA@CM-AP, MSNs@LA@CM-AP or MSNs@RA/LA@CM-AP (Fig. 3A-D). Consistently, these formulations reduced collagen-1a2 gene expression and collagen secretion of aHSCs (Fig. 3E and F). Additionally, MSNs@RA@CM-AP showed better antifibrotic effects than free RA due to the improved cellular internalization of RA by the nanocarrier system. Notably, compared with MSNs@RA@CM-AP, MSNs@RA/LA@CM-AP induced lower expression of fibrogenic markers and less secretion of collagen, which could be explained by the fact that norepinephrine is an autocrine factor for HSCs and that the loaded LA inhibited the self-produced norepinephrine from promoting the proliferation and activation of HSCs through the autocrine closed loop.
The excessive production of ROS plays a vital role in activating the HSCs to generate abundant collagen during liver injuries. We expected that our MSNs could scavenge excessive ROS to inhibit HSC activation. As shown in Fig. 3G and H, LPS stimulation significantly increased the intravenous ROS level of LX-2 cells. However, such effect was obviously inhibited by the treatments of MSNs@CM-AP and MSNs@RA/LA@CM-AP. Additionally, both MSNs@CM-AP and MSNs@RA/LA@CM-AP suppressed the increasement of α-SMA and collagen-1a2 expression in LPS-activated LX-2 cells (Fig. 3I and J). Consistent results were detected for the secretion of proinflammatory cytokines including TNF-α, IL-1β and IL-6 in LPS-stimulated LX-2 cells (Fig. 3K-M). These results indicated the anti-oxidant and anti-inflammatory capabilities of MSNs@RA/LA@CM-AP, which might benefit the NAFLD treatment.
To investigate the cellular crosstalk between HSCs and sympathetic neurons, we constructed a coculture system of aHSCs and sympathetic neurons in a Transwell model. As illustrated in Fig. 4A, the number of HSCs was significantly greater after coculture with sympathetic neurons for 72 h than after coculture with free DMEM. Furthermore, increased expression of α-SMA, Col1a1, Col3a1, Col5a1 and collagen-1a2 and increased secretion of collagen were detected in HSCs cocultured with sympathetic neurons (Fig. 4B-E and S10). Additionally, the addition of NE to HSCs cultured with free DMEM also induced an increase in cell viability and fibrosis-related gene expression in HSCs. To further explore the bidirectional influence, we detected the production of sympathetic nerve neurotransmitters in the coculture system. The coculture system secreted a surge of norepinephrine, which was markedly greater than that secreted by single-cultured sympathetic neurons (Figure S11), suggesting that sympathetic nerves were activated by HSCs. These results suggested that sympathetic neurons increased the proliferation and activation of HSCs. We expected that the undesirable crosstalk between HSCs and sympathetic neurons could be inhibited by the nanoplatform to achieve efficient treatment of hepatic fibrosis. Consistent with our expectation, the proliferation and activation of HSCs were significantly inhibited by the administration of MSNs@LA@CM-AP, MSNs@LA@CM-AP and MSNs@RA/LA@CM-AP (Fig. 4A-E and S10). Moreover, MSNs@RA/LA@CM-AP showed a greater deactivation effect than MSNs@RA@CM-AP and MSNs@LA@CM-AP, and strongly suppressed the proliferation and activation of HSCs after the addition of NE. These results indicated that blockade of the adrenoceptor could improve the HSC deactivation effect of the nanoplatform.
To explore the lipid-lowering effect of MSNs@RA/LA@CM-AP, TG and TC levels in FFA-treated HepG2 cells were detected after various treatments. FFA treatment indeed elevated the TG and TC contents in HepG2 cells. However, MSNs@RA@CM-AP induced significant decreases in TG and TC contents similar to MSNs@RA/LA@CM-AP, which was likely because that RA inhibited FFA uptake and lipogenesis (Figure S12). To investigate whether MSNs@RA/LA@CM-AP could decrease cellular lipid deposition by interrupting crosstalk between hepatocytes and sympathetic nerves, sympathetic neurons seeded on the upper chamber and FFA-treated HepG2 cells were layered on the bottom chamber with various formulations. Figure 4G and S13 indicates that the TG and TC levels were further increased in the FFA-treated HepG2 cells when cocultured with sympathetic neurons. However, the upregulation of TG and TC induced by sympathetic neurons was completely eliminated by MSNs@LA@CM-AP. Notably, MSNs@RA/LA@CM-AP led to the greatest reduction in lipid deposition in FFA-treated HepG2 cells. Consistently, Bodipy staining revealed that the lipid deposition was obviously lower in steatotic HepG2 cells after treatment with MSNs@RA/LA@CM-AP than those treatments with MSNs@RA @CM-AP and MSNs@LA@CM-AP (Fig. 4H and S14). These results suggested that MSNs@RA/LA@CM-AP integrating adrenoceptor antagonist had a beneficial lipid-lowering effect.
We investigated the antifibrotic effect of MSNs@RA/LA@CM-AP in vivo using BDL-induced liver fibrosis models. As shown in Fig. 5A-C and S15, MSNs@CM-AP slightly reduced the α-SMA expression and collagen deposition as well as pathological area in the liver tissue, which was possibly due to the ROS-scavenging effect of MSNs. Furthermore, both MSNs@RA@CM-AP and MSNs@LA@CM-AP partly alleviated liver fibrosis. Compared with treatment with MSNs@RA@CM-AP and MSNs@LA@CM-AP, treatment with MSNs@RA/LA@CM-AP led to a greater decrease in α-SMA expression and collagen deposition, indicating the potential benefits of the blockade of adrenoceptors in the nanoplatform on HSC inactivation. Additionally, MSNs@RA@CM-AP, MSNs@LA@CM-AP and MSNs@RA/LA@CM-AP elicited a striking reduction in liver parenchymal necrosis and the liver fibrotic area compared with saline and MSNs@CM-AP. Furthermore, ALT and AST, the main biochemical markers of liver function, as well as hydroxyproline, an important component of the ECM that reflects collagen deposition and the severity of liver fibrosis, significantly decreased in the MSNs@RA@CM-AP, MSNs@LA@CM-AP and MSNs@RA/LA@CM-AP groups (Fig. 5D-F). Notably, the ability of MSNs@RA/LA@CM-AP to reduce liver fibrosis and damage was greater than that of MSNs@RA@CM-AP and MSNs@LA@CM-AP. These results further confirmed that the combination of MSNs@RA/LA@CM-AP with adrenoceptor antagonists could enhance the treatment of liver fibrosis.
NAFLD is considered a result of liver steatosis. The activation of liver sympathetic nerves regulates liver steatosis during obesity. To explore whether MSNs@RA/LA@CM-AP combined with adrenoceptor antagonists could decrease obesity-induced liver steatosis to ameliorate NAFLD, NAFLD mouse models were established through long-term high-fat diet (HFD) feeding, and these mice were subsequently administered MSNs@RA/LA@CM-AP or other formulations (Fig. 6A). As anticipated, the body weight and liver weight were greater in the mice after 10 weeks of HFD feeding than those in the mice fed the normal diet (healthy mice) (Fig. 6B and C). Additionally, the sympathetic nerves of the HFD-fed mice exhibited increased activity compared with those of the normal chow-fed mice, as evidenced by the increased production of norepinephrine (Figure S16). These findings further indicated that HFD-induced NAFLD is associated with liver sympathetic nerve overactivity. Notably, all treatments, especially MSNs@RA/LA@CM-AP, significantly decreased the body weight, liver weight and liver/body weight ratio of HFD-fed mice (Fig. 6B and D). Liver lipid deposition is an important issue in NAFLD; therefore, lipid levels were detected to investigate the lipid-lowering efficacy of MSNs@RA/LA@CM-AP. The levels of TC, TG and LDL-C in the liver and serum of mice after treatment with MSNs@RA/LA@CM-AP were lower than those in the liver and serum of mice after treatment with MSNs@RA@CM-AP and MSNs@LA@CM-AP (Fig. 6E-F and Figure S17). Additionally, oil red O staining of neutral lipids revealed that MSNs@RA/LA@CM-AP led to the least lipid droplet accumulation in the cytoplasm of liver tissues, indicating that MSNs@RA/LA@CM-AP exerted a lipid-lowering effect (Fig. 6I). Furthermore, the ALT and AST levels in the blood were the lowest in the MSNs@RA/LA@CM-AP group among other treatment groups in the NAFLD mouse models, which were approximate to those in normal mice (Fig. 6G and H). These results confirmed the excellent ability of MSNs@RA/LA@CM-AP to reduce liver steatosis and damage, suggesting its tremendous potential in NAFLD therapy.
NAFLD is a chronic disease associated with steatosis, inflammation and fibrosis. Effective treatments of NAFLD remained a great challenge due to the limited available therapeutic options in the clinic. Increasing evidence has revealed that hepatic sympathetic innervation promoted the activation and proliferation of HSC and regulated liver lipid acquisition during obesity through adrenoceptors expressed by HSCs and hepatocytes. Therefore, adrenoceptors might be promising targets for novel approaches to treat NAFLD.In the current study, we hypothesized that blockade of adrenoceptors for breaking the vicious crosstalk of sympathetic nerves in the liver might provide benefits to the treatment of NAFLD. Herein, a biomimetic nanocarrier was tailored to co-deliver RA, a commonly used anti-NAFLD agent and LA, an adrenoceptor antagonist. The biomimetic nanocarrier itself consumed the excessive ROS at the pathologic site, which contributed to decrease the inflammatory response and inhibit the fibrosis progression. Furthermore, the biomimetic nanocarrier possessed aHSC-targeting property and ROS-responsiveness release as well as Kupffer cell-evading ability, which facilitated the drug delivery and improved the therapeutic efficiency of RA for NAFLD.
Norepinephrine, as an autocrine factor produced from HSCs, can also promote the proliferation of HSCs. The MSNs@RA/LA@CM-AP integrating the blockade of adrenoceptors effectively inhibit the stimulatory effects of self-produced norepinephrine on the HSC proliferation. Importantly, we found that our prepared MSNs@RA/LA@CM-AP intercepted cellular crosstalk of sympathetic neurons with HSCs, which effectively inhibited the activated and proliferative effect of sympathetic neurons on HSCs. Therefore, MSNs@RA/LA@CM-AP showed great ability to reduce liver fibrosis and damage in BDL-induced liver fibrosis animal models
There are no obvious differences in the lipid accumulation of steatotic HepG2 cells after treated with MSNs@RA@CM-AP and MSNs@RA/LA@CM-AP, which was possibly because the lipid-lowering effect was mainly originated from the reduction of FFA uptake and lipogenesis by RA[20]. However, MSNs@RA/LA@CM-AP exhibited better lipid-lowering effect in steatotic HepG2 cocultured with sympathetic neurons and in HFD-induced NAFLD animal models. Norepinephrine can act on the overexpression of adrenoceptors in hepatocytes, resulting in an increase in lipid deposition according to the previous report[24]. Therefore, MSNs@RA/LA@CM-AP integrating adrenoceptor antagonist exhibited excellent ability to reduce liver steatosis, which further indicated that our nanoplatform regulated all stages of NAFLD involving steatosis, inflammation and fibrosis.
{kind=link}