GPC, grain size, and spike density increased in S-Cp2
The agronomic traits and processing quality parameters of S-Cp2 and its WT control were evaluated in the 2021–2022 and 2022–2023 growing seasons. The comparison with the WT control revealed grain length, grain width, TKW, and spike density were higher in S-Cp2, whereas the opposite trend was observed for volume weight, plant height, spike length, spikelet number per spike, and grain number per spike (Fig. 1; Tables 1 and 2).
Table 1
Comparison of the agronomic traits of S-Cp2 and the WT control
| Traits | Growing season | WT | S-Cp2 | E | G | E × G |
| Plant height (cm) | 2021‒2022 | 72.30 ± 1.51 | 49.43 ± 1.96** | 2365.86** | 6423.19** | 85.30** |
| 2022‒2023 | 82.39 ± 1.92 | 64.24 ± 1.81** |
| Spike length (cm) | 2021‒2022 | 12.37 ± 0.91 | 6.53 ± 0.50** | 0.03 | 4267.80** | 10.78** |
| 2022‒2023 | 12.69 ± 0.74 | 6.24 ± 0.37** |
| Spikelet number per spike | 2021‒2022 | 21.26 ± 0.90 | 20.74 ± 1.26* | 36.40** | 23.63** | 3.30 |
| 2022‒2023 | 22.60 ± 1.09 | 21.46 ± 1.50** |
| Spike density | 2021‒2022 | 1.72 ± 0.10 | 3.19 ± 0.25** | 276.80** | 58.78** | 37.90** |
| 2022‒2023 | 1.78 ± 0.09 | 3.44 ± 0.15** |
| Grain number per spike | 2021‒2022 | 58.02 ± 5.88 | 44.70 ± 2.40** | 361.21** | 1366.07** | 132.66** |
| 2022‒2023 | 54.10 ± 2.91 | 28.72 ± 2.45** |
| Grain length (cm) | 2021‒2022 | 7.34 ± 0.22 | 7.48 ± 0.24* | 18.19** | 26.27** | 0.61 |
| 2022–2023 | 7.45 ± 0.20 | 7.64 ± 0.22** |
| Grain width (cm) | 2021‒2022 | 3.29 ± 0.12 | 3.69 ± 0.15** | 23.84** | 192.26** | 19.59** |
| 2022‒2023 | 3.49 ± 0.12 | 3.70 ± 0.21** |
| Leaf length (cm) | 2021‒2022 | 18.17 ± 1.65 | 21.91 ± 2.70* | 6725.31** | 55.58** | 84.86** |
| 2022‒2023 | 19.65 ± 2.05 | 22.35 ± 2.37** |
| Leaf width (cm) | 2021‒2022 | 2.12 ± 0.14 | 1.65 ± 0.13** | 1580.20** | 1051.49** | 3407.83** |
| 2022‒2023 | 1.85 ± 0.18 | 1.45 ± 0.11** |
| Thousand kernel weight (g) | 2021‒2022 | 50.77 ± 0.79 | 55.97 ± 0.60** | 10.02** | 440.06** | 9.46** |
| 2022‒2023 | 50.80 ± 0.46 | 57.79 ± 0.70** |
| Productive tiller number | 2021‒2022 | 3.86 ± 0.86 | 3.98 ± 0.94 | 5112.06** | 34.84** | 29.16** |
| 2022‒2023 | 4.14 ± 1.03 | 4.30 ± 1.05 |
| **, P < 0.01; *, P < 0.05; E, environment; G, genotype; E × G, interaction between the environment and genotype. Data are presented as the mean ± standard deviation. |
Table 2
Comparison of the processing quality parameters of S-Cp2 and the WT control
| Traits | Growing season season | WT | S-Cp2 | E | G | E × G |
| Grain protein content (%, dry weight) | 2021‒2022 | 12.50 ± 0.37 | 14.81 ± 0.38** | 354.72** | 142.60** | 1.19 |
| 2022‒2023 | 16.27 ± 0.55 | 19.05 ± 0.57** |
| Zeleny sedimentation value (mL) | 2021‒2022 | 14.22 ± 0.53 | 17.41 ± 0.40** | 1127.91** | 87.79** | 1.05 |
| 2022‒2023 | 24.84 ± 0.83 | 27.40 ± 0.87** |
| Wet gluten content (%) | 2021‒2022 | 20.77 ± 1.10 | 25.20 ± 1.04** | 689.96** | 149.31** | 8.90** |
| 2022‒2023 | 31.95 ± 0.86 | 39.25 ± 1.25** |
| Water absorption (%) | 2021‒2022 | 55.62 ± 0.46 | 59.12 ± 0.83** | 2.08 | 190.10** | 9.24** |
| 2022‒2023 | 55.10 ± 0.67 | 60.58 ± 0.88** |
| Development time (min) | 2021‒2022 | 1.49 ± 0.08 | 2.03 ± 0.09** | 173.17** | 294.95** | 72.92** |
| 2022‒2023 | 1.78 ± 0.16 | 3.40 ± 0.20** |
| Volume weight (g) | 2021‒2022 | 799.90 ± 5.85 | 784.90 ± 3.96** | 17.88** | 244.15** | 52.97** |
| 2022‒2023 | 805.38 ± 3.01 | 764.22 ± 2.37** |
| Loaf height (mm) | 2021‒2022 | 54.20 ± 2.59 | 64.00 ± 4.18** | 2.06 | 49.69** | 0.88 |
| 2022‒2023 | 55.00 ± 4.00 | 67.80 ± 3.35** |
| Loaf volume (mL) | 2021‒2022 | 166.00 ± 13.04 | 198.40 ± 16.44** | 0.91 | 31.87** | 0.11 |
| 2022‒2023 | 169.80 ± 12.21 | 206.20 ± 12.38** |
| **, P < 0.01; *, P < 0.05; E, environment; G, genotype; E × G, interaction between the environment and genotype. Data are presented as the mean ± standard deviation. |
To determine whether Gpc-5B affects wheat processing quality, quality-related parameters were compared between S-Cp2 and the WT control. In addition to GPC, the gliadin, high-molecular-weight glutenin subunit (HMW-GS), low-molecular-weight glutenin subunit (LMW-GS), and glutenin contents were significantly higher for S-Cp2 than for the WT control (Fig. 2c). As expected, the Zeleny sedimentation value, wet gluten content, development time, water absorption, and loaf volume were higher for S-Cp2 than for the WT control (Fig. 2a, b; Table 2).
Mutant phenotype of S-Cp2 was controlled by a single dominant nuclear locus
S-Cp2 was reciprocally backcrossed with ‘Shumai482’ to clarify the genetic basis of the mutant phenotype. All BC1F1 plants had a compact spike, suggesting that the mutant phenotype was controlled by at least one nuclear gene. The mutant traits of S-Cp2, especially the increases in GPC, grain size, and spike density, were not separated in the BC1F2 population (S-Cp2 × ‘Shumai482’; 400 individuals), implying these traits were controlled by the same locus. To facilitate genetic research, compact spike was selected as the target trait for the subsequent χ2 test. S-Cp2 was crossed with three common wheat lines (i.e., ‘Br196’, ‘Mianmai112’, and ‘Shumai133’). In the S-Cp2 × ‘Br196’ F2 population, 134 plants had a compact spike and 46 plants had a normal spike, which was consistent with the theoretical 3:1 segregation ratio (χ2 = 0.03 < χ20.05 = 3.84). Similarly, the corresponding ratios in the S-Cp2 × ‘Mianmai112’ F2 population (207 plants with a compact spike and 73 plants with a normal spike; χ2 = 0.17 < 3.84) and the S-Cp2 × ‘Shumai133’ F2 population (174 plants with a compact spike and 55 plants with a normal spike; χ2 = 0.12 < 3.84) also fit the expected 3:1 segregation ratio. These results suggest the mutant phenotype of S-Cp2 was controlled by a single dominant nuclear locus (Gpc-5B).
Mapping of Gpc-5B
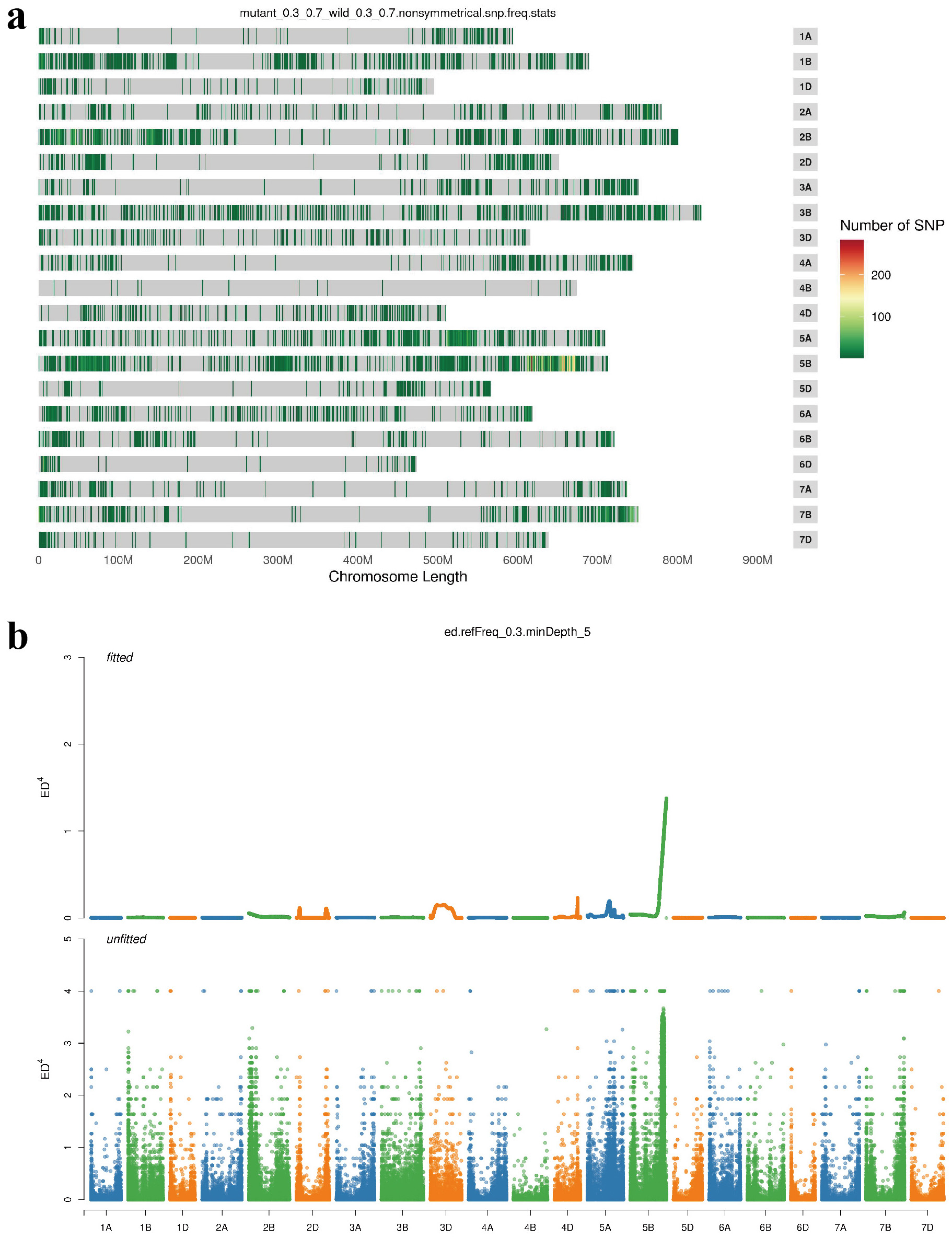
The F2 population derived from the S-Cp2 × ‘Br196’ cross was used to map Gpc-5B according to the BSE-seq analysis. After filtering for read-depth quality, 16,900 SNPs between the wild bulk and mutant bulk were concentrated in the 655–665 Mb region of wheat chromosome 5B (Fig. S1a). The analysis of Euclidean distance revealed a continuous distribution peak on chromosome 5BL (Fig. S1b), indicating that Gpc-5B was located on 5BL. Using KASP markers (Table S2), Gpc-5B was mapped between markers KASP5771 and KASP6597, with a genetic distance of 1.5 cM (Fig. 3a, b).
To more precisely map Gpc-5B, 130 F2 heterozygous plants from the S-Cp2 × ‘Br196’ cross were self-pollinated to generate an F3 population consisting of 2,450 individuals. Six newly developed KASP markers (Table S2) were used for screening recombinants (RM1–RM6). Finally, Gpc-5B was delimited to an 87.24 kb interval between markers KASP58070 and KASP58158 (Fig. 3c, d). This region included three high-confidence predicted genes (TraesCS5B02G486800, TraesCS5B02G486900, and TraesCS5B02G487000; Fig. 3e).
5 Bq is the gene underlying the Gpc-5B locus
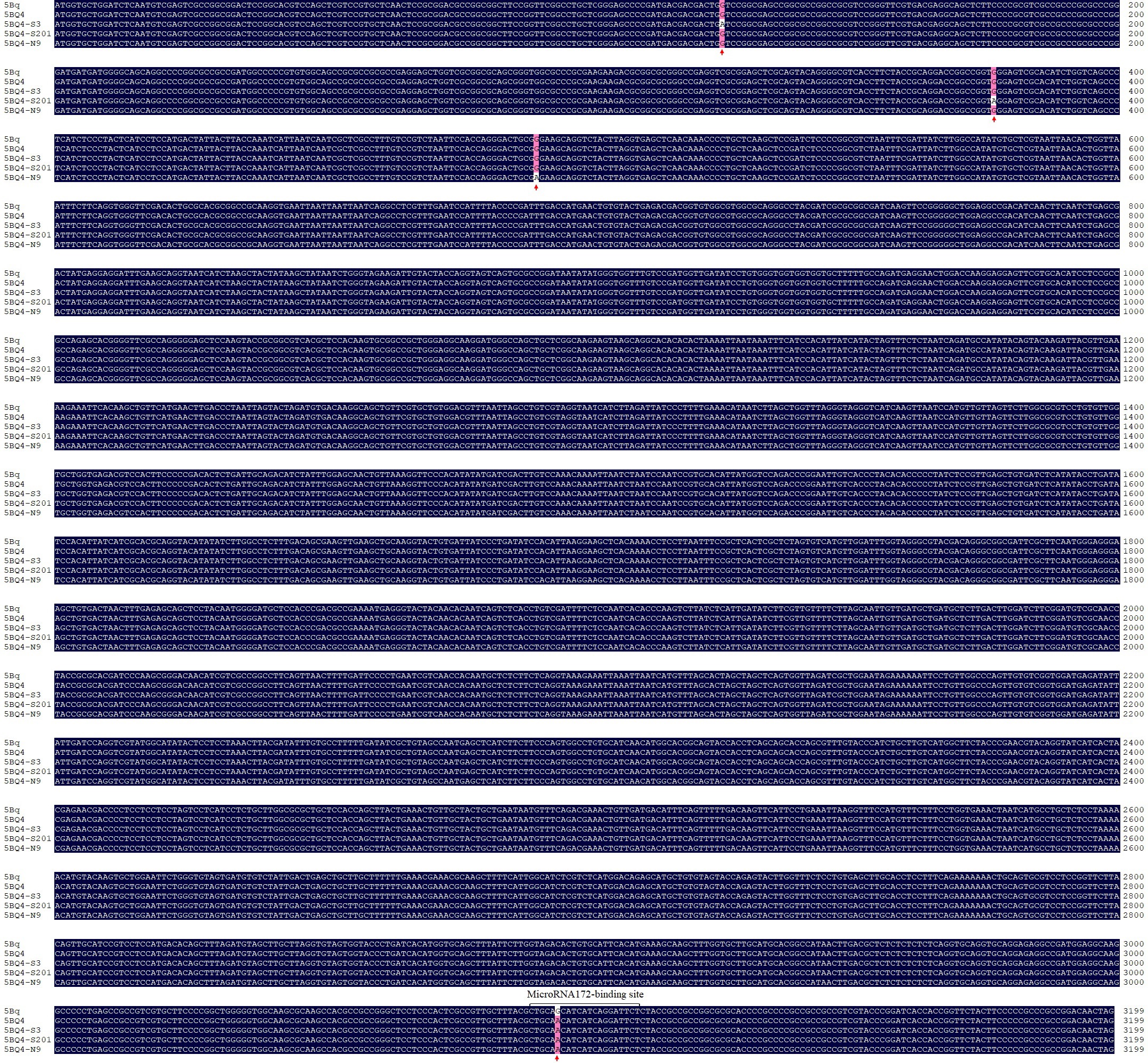
Among the three high-confidence predicted genes, TraesCS5B02G486900 (5Bq), which is homologous to the Q gene on 5AL, was hypothesized to be the candidate gene for Gpc-5B. To test this hypothesis, 5Bq cDNA and genomic sequences were cloned for both S-Cp2 and ‘Shumai482’. A missense mutation (G-to-A) was detected in the microRNA172-binding site of the 5Bq coding sequence in S-Cp2 (Figs S2–S4), resulting in an amino acid change from alanine to threonine. This mutant 5Bq allele in S-Cp2 was named 5BQ4 (GenBank No. PP584578). A qRT-PCR analysis of the roots, stems, and leaves indicated 5BQ4 transcript levels in S-Cp2 were significantly higher (P < 0.01) than 5Bq transcript levels in ‘Shumai482’ (Fig. 2d).
To further assess whether 5Bq is the gene responsible for the mutant phenotype, S-Cp2 seeds were treated with EMS to obtain revertants, among which three (S-Cp2-rev1, S-Cp2-rev2, and S-Cp2-rev3) had a normal spike (Fig. 4). The mutant 5Bq alleles in S-Cp2-rev1 (nonsense mutation revealed by the comparison with 5BQ4), S-Cp2-rev2 (nonsense mutation), and S-Cp2-rev3 (missense mutation) were designated as 5BQ4-S3 (GenBank No. PP584579), 5BQ4-S201 (GenBank No. PP584581), and 5BQ4-N9 (GenBank No. PP584580), respectively (Figs S2–S4).
Haplotype analysis
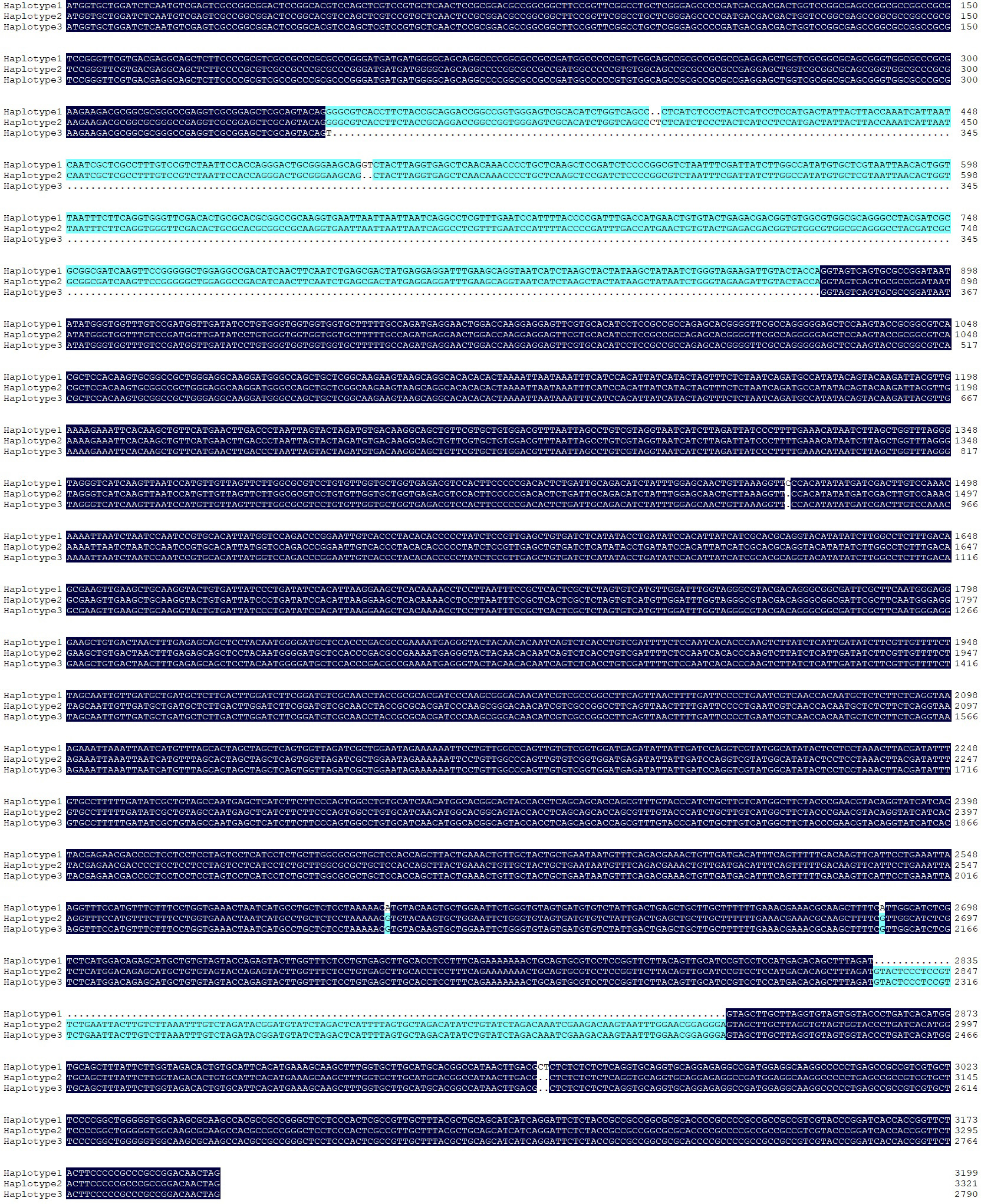
To clarify haplotype variations, 5Bq coding sequences in 30 common wheat accessions were cloned and analyzed (Table S1). Three haplotypes (haplotypes 1 [functional], 2 [pseudogene], and 3 [pseudogene]) were identified (Figs S5 and S6). Two specific markers (5BqDNA-1 and 5BqDNA-3) were developed to distinguish haplotypes (Fig. 5). We performed a haplotype analysis of 5Bq in 210 landraces and 190 cultivars (Table S1). Notably, the proportion of functional haplotype 1 increased from 4.76% in landraces to 39.48% in modern cultivars.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}