Dosage dependence effect of CNP on the rice seedlings’ phenotypic growth
CNP had induced several positive growth effects in rice plants (Panigrahy et al., 2021). Hence, it was necessary to determine the ineffective, optimum and maximum concentrations of CNP to show positive growth effects. Study of seedling shoot length (SL), root length (RL), rootlet number (RLN) and nodal roots (NR) at 7 different CNP concentrations (ranging from 200 µg/ml to 1500 µg/ml) (Fig. 1, Suppl Fig. 1) showed that SL, RL and RLN tend to increase after at least 350 µg/ml indicating positive growth effects. SL and RL showed highest increase between 500 and 750 µg/ml after which they started to decline (Fig. 1A). RLN also was highest number at 750 µg/ml (Fig. 1).
These results indicated that the optimum concentration of CNP on rice seedlings were between 500 and 750 µg/ml. NR showed opposite trend with in increase after at least 750 µg/ml and was higher till 1500 µg/ml of CNP (Fig. 1). This further indicated that NR could be a toxicity response induced due to CNP between 1000–1500 µg/ml. Germination percentage on 4th day of rice seeds was homogenously 100% in all concentration indicating that CNP didn’t exert any effect on seed germination or growth of seedlings.
Effect of CNP treatment on physiological parameters in rice plants
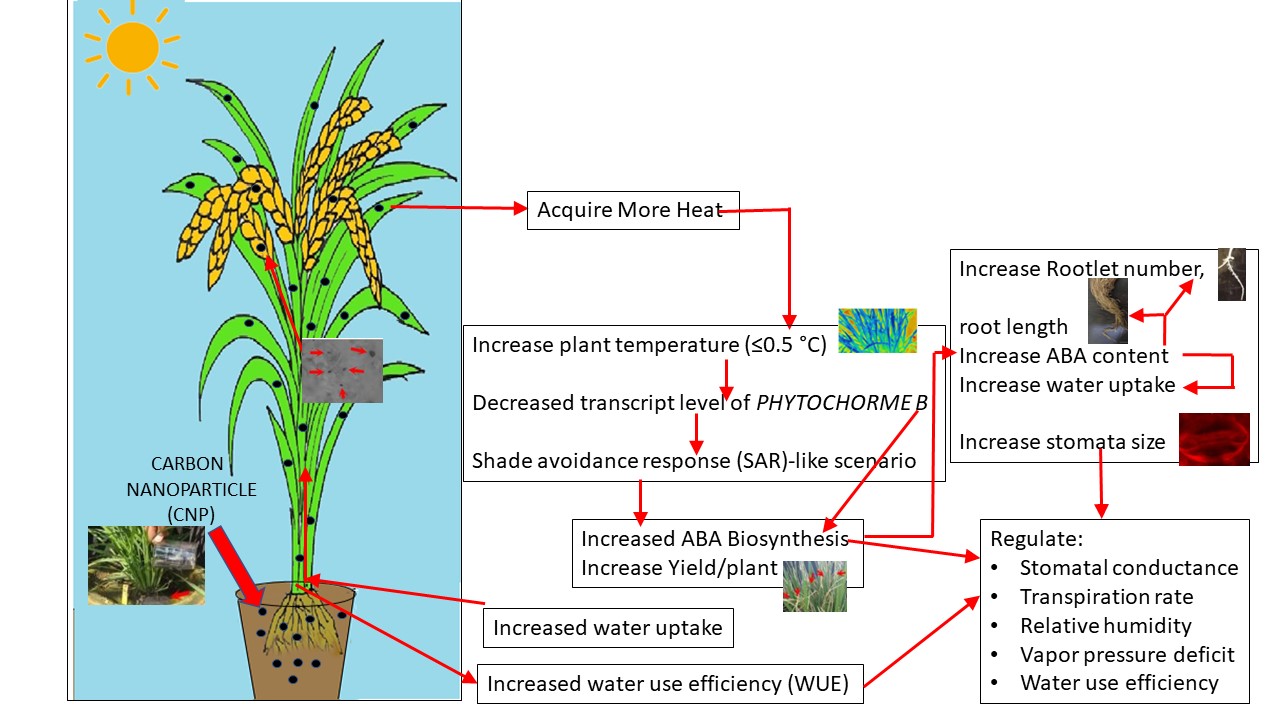
Water relation parameters were assessed after four CNP treatments in the treated and untreated plants to investigate the reason of improved growth and yield of CNP-treated plants (Fig. 1B-G)). Photosynthesis parameters such as photosynthetically active radiation inside the leaf (PARi) (Fig. 1D), carbon assimilation (A) (Fig. 1F) showed significantly higher values in the CNP-treated plants. Water relation parameters such as leaf temperature (Tleaf) (Fig. 1B), and water use efficiency (WUE) (Fig. 1G) were significantly higher (by 0.4°C ± 0.1°C and 75% respectively) in the CNP-treated plants. Relative humidity (RH) (Fig. 1C) was significantly less (i.e. 13.3%) in CNP-treated plants. Vapour pressure deficit (VPD) (Fig. 2F) and transpiration rate (E) showed negligible changes between the CNP-treated and the control plants (data not shown). Stomatal conductance was also found to be higher in the CNP-treated plants (Fig. 1E). CIRASS3 measurements and analysis showed improvements in the water relation traits in the CNP treated plants as compared to their controls.
Effect of CNP treatment on stomatal parameters in rice plants
As the CNP-treated plants had improved water relation parameters, stomatal analysis was performed using confocal microscopy in the control and CNP-treated samples in the seedlings stage as well as after 4th CNP treatment in plants (Fig. 2).
Stomata in the seedling cells showed clear bulging in the cell wall and distortions of the CNP-treated samples which were completely absent in the control samples (Fig. 2A-2C). Samples from older plants also showed similar bulging and distortions (Fig. 2D-2F). Frequency of stomata per unit area of leaf decreased significantly (~ 50%) in both the seedlings and older plant leaves (Fig. 2G) with an increase of ~ 6% in the size of the stomata (Fig. 2H). These results indicated that CNP treatment might have affected to increase stomatal size and decrease in their frequency.
Analysis of mechanism of CNP uptake
Water uptake by the seedlings with or without CNP in the medium was quantified studying the % increase or decrease of water uptake per seedling (Fig. 3).
Addition of CNP in the medium increased water uptake (i.e., 13%) by the seedlings (Fig. 3I). To understand if CNP uptake is coupled to water uptake or through exo-/ endo- or pinocytosis, inhibitor nocodazole was added with or without CNP in the medium. Treatment with nocodazole increased the water uptake by 16% than the control with water alone (Fig. 3I). Nocodazole treatment in the CNP-treated seedlings reduced the water uptake (6% more than control water alone) as compared to those CNP treated ones (Fig. 3I). To investigate the reason of increased water uptake with CNP, number of developed xylem strands and their average perimeter was measured from confocal microscopy images in CNP-treated root samples. Control seedlings with water alone showed 13 ± 1 developed xylem strands with an average perimeter of ~ 11.6 µm (Fig. 3A, 3E, 3K). Seedlings grown with nocodazole showed reduced number (i.e., 8 ± 1) of developed xylem strands but no change in average perimeter (Fig. 3B, 3F, 3K). CNP-treated seedlings showed increase in number of developed xylems (i.e., 14–15 ± 1), which were found in 2–3 rings also with increased average perimeter of developed xylem strands of ~ 15 µm (Fig. 3C, 3G, 3K). Inhibitor application with CNP treatment showed similar number of developed xylem strands as that of CNP, however the average diameter was reduced to ~ 10.9 µm indicating reduced water uptake ((Fig. 3D, 3H, 3K). Further, an estimate of CNP uptake was made by measuring the area of CNP aggregates in the confocal microscopy images. CNP aggregates in the nocodazole treated samples similar to that with the CNP-treatment alone (Fig. 3J).
Genome-wide expression and pathway analysis of transcripts during CNP-treatment
Our first report in Arabidopsis thaliana (A.thaliana; At) plants with CNP treatment induced accelerated flowering, photoperiod dependency (Kumar et al., 2018) and in our second study on rice, CNP treatment resulted improved growth and yield along with shade avoidance response (SAR) phenotype and increased plant temperature, albeit with minimal preponement of flowering time (Panigrahy et al., 2021). To understand the basis of the differences in CNP effects in the model dicot and monocot plants, transcriptome analysis and microarray experiment was performed separately from seedling or leaf samples of A. thaliana and flag leaf samples of Swarna rice variety respectively (Fig. 4).
Each of these experiments were done with 2 biological replicates from control condition or with CNP. At transcriptome volcano plot for transcripts below 1.0 (Fig. 4A) showed that highest number of transcripts (37.4%) had p < 0.5. Moreover, the replicates of control libraries or the test libraries bear more collinearity (Fig. 4C), making them suitable platform for further analysis. Standard analysis of the transcripts (described in Methods section) resulted 23098 differentially expressed transcripts (DEG) which were found in both with and without CNP (Figure. 4B). Pathway enrichment analysis showed the fold-enrichment of the Up- or Down-regulated DEGs (Fig. 4D, 4E) with the number of genes of the top 30 pathways. Among the up-regulated DEGs, ‘Plant hormone signal transduction pathways’ was highly enriched with fold enrichment 3.78, enrichment FDR value 3.77413E-11 containing 40 genes in list (Fig. 4D). And among the down-DEGs, unsaturated fatty acids such as ‘α-linolenic acid metabolism’ (fold enrichment 4.79, enrichment FDR 1.68024E-05) followed by ‘phenylpropanoid biosynthesis’ with fold enrichment of 2.52 and enrichment FDR of 5.02073E-05 were prominent (Fig. 4E). These results indicated that CNP treatment in A.thaliana alleviated lipids and secondary metabolites with decreased alpha-linolenic acid like stress antioxidants. Moreover, it indicated an increased involvement of hormone signal transduction. Pathway category analysis of At transcriptome DEGs showed that abscisic acid (ABA) pathway genes were involved to the highest followed by the auxin pathway genes (Figure. 4F). Microarray analysis from CNP-treated and untreated samples of Swarna rice flag leaf samples resulted 52–59% of detection of probes in the control and test arrays, with 28,482 probes detected in common (Fig. 4G). Collinearity among the control and test replicates above 0.8 (Figure. 4H) created suitable platform for further analysis. Function and pathway analysis showed that major category of transcripts in the CNP-treated samples were from cell membrane, DNA or RNA, transcriptional activity, phosphate homeostasis genes. Hormone pathway genes acquired 2.53% of the total genes involved (Fig. 4I). Cohering to the pathway result of A.thaliana, pathway analysis of rice samples revealed that ABA pathway genes were highly enriched being the second most enriched after auxin pathway genes (Figure. 4J). Out of the 34 most prominent ABA pathway genes, 29 were found to be up-regulated in CNP-treated samples (Figure. 4K).
Comparative study of functional categories of differentially expressed genes (DEGs) among rice and Arabidopsis
DEGs from the At transcriptome and rice (Os) microarray were arranged according to their function (GO annotation), pathway involved (KEGG database) and details obtained from TAIR and RAP-annotations for Arabidopsis and rice respectively. These relevant categories included ABA pathway, cold stress response, light signaling and circadian clock, photosynthesis and carbohydrate, ROS detoxification, ion and water transport, stomata size, which were used for the comparative analysis among the two model plant systems (Table S1, Table S2). In ABA pathway, 34 genes were found among which majority (24 genes) were up-regulated in rice. Whereas, in Arabidopsis only 12 gene were found to be involved in ABA pathway, most of which (~ 66%) are down-regulated. Cold stress response category of genes represented different regulation among At and Os. In rice ~ 54% of these genes were up-regulated, whereas in At most of the genes in this category were down-regulated. Genes of light signaling, and circadian clock didn’t represent any significant differences in their numbers of up and down DEGs among rice and At. In photosynthesis, chlorophyll and carbohydrate category, all the DEGs were down-regulated in rice while ~ 63% of DEGs were up-regulated in At. DEGs of ROS detoxification category presented contrasting pattern with majority being down-regulated in At and up-regulated in rice. However, the transcript levels of superoxide dismutase and catalase in At data were upregulated (Table S2). Also, the 11 class III Peroxidases found in rice microarray data were all up-regulated indicating these may act to detoxify the ROS in CNP-treated samples. The DEGs of ion, solute and water transport category were synergistically presented being majority of them down-regulated in both At and rice. DEGs of stomata size were up-regulated in both At and rice. These results indicated that CNP treatment induced differential effect on regulation of genes of different functional categories in At or rice.
Effect of CNP treatment on endogenous Abscisic acid content and synthesis
To understand if CNP treatment with abscisic acid (ABA) has any effect on seedling root and shoot growth, (ABA) was added to the growth medium to observe its effect on seedling phenotype. ABA is known to slow down the root and shoot growth in rice (Chen et al., 2006). ABA treatment without or with CNP resulted comparable % of decrease in SL and RL respectively (Fig. 5A).
While SL decreased 74.6% and 72.2%, RL decreased 77.2% and 71.6% in case of control and CNP-treated seedlings respectively (Fig. 5A). RLN in the CNP-treated seedlings decreased to nearly half, however surprisingly, the control seedlings showed significant increase of ~ 2.2-fold in the RLN with ABA treatment (Fig. 5A). These results indicated that CNP induced growth effects could alter endogenous ABA levels. Endogenous ABA in seedlings showed that CNP-treated seedlings had nearly double amount of endogenous ABA than the untreated ones (Fig. 5B). Congruently, flag leaf samples of CNP-treated plants were found to have ~ 40-fold higher ABA content than the untreated ones (Fig. 5C). Additionally, the three other defense hormones such as jasmonic acid (JA), salicylic acid (SA) and oxylipins (oxy) were also found to be higher in the CNP-treated plants (Suppl. Figure 2). Transcript expression analysis of ABA biosynthesis gene 9-cis-epoxycarotenoid dioxygenase 5 (AtNCED6) in Arabidopsis seedlings (Fig. 5D) and 9-cis-epoxycarotenoid dioxygenase 1 (OsNCED1) in rice flag leaves (Fig. 5E) were validated to support the above observation of ABA content. AtNCED6 relative expression level was increased to ~ 18-fold and OsNCED1 relative expression level was decreased to ~ 0.39-fold after CNP treatment.
Transcript expression analysis to understand CNP-treated plants’ phenotype
To have insights into CNP-treated plant’s phenotype, transcript expression of different selected genes from photosynthesis, calvin cycle, cold tolerance, stomata size and frequency, genes for ABA pathway were studied using qPCR in CNP-treated plants in comparison with their control plants. The concordance of the transcript expression levels of majority of the genes analysed from the rice microarray with the qRT-PCR fold change provided evidence of sound genomic data set (Suppl. Figure 3A). Photosynthesis related gene OXYGEN EVOLVING COMPLEX1 (OsOEE1) was significantly down-regulated to 0.25-fold in CNP-treated samples compared to the controls (Fig. 6A).
SEDOHEPTULOSE 1-7-BI-PHOSPHATE (OsSBPase) (Suzuki et al., 2021, Feng et al., 2007) was significantly down-regulated to 0.27-folds in CNP-treated samples compared to their controls (Fig. 6B). These results indicated that the input to the calvin cycle through the OsSBPase as well as the photosynthetic yield, as indicated through OsOEE1 (Heide et al 2004) transcript expression levels, were deteriorated in the CNP-treated plants compared to the untreated plants. To confirm the microscopic observations of stomatal parameters in the CNP-treated plants, genes related to stomatal size and frequency including EPIDERMAL PATTERNING FACTOR-LIKE 9 (OsEPFL9) and SUCROSE NON-FERMENTING-1 RELATED PROTEIN KINASE 2 (OsSNRK2), were studied for their transcript expression levels. OsEPFL9 transcript expression level was found to be reduced to ~ 0.31-folds in the CNP-treated samples compared to the controls (Fig. 6C). OsSNRK2 transcript expression level in the CNP-treated plants was found to be reduced to nearly half (~ 0.55-fold) compared to their controls (Fig. 6D). To understand the molecular basis of the altered water relation parameters in the CNP-treated plants, water use efficiency (WUE) related aquaporin gene PLASMA MEMBRANE INTRINSIC PROTEIN 2–5 (OsPIP2-5) transcript level was studied comparatively. OsPIP2-5 transcript level was significantly increased to ~ 2.5-fold in the CNP-treated seedlings compared to the controls (Fig. 6E). To confirm the involvement of CNP in abiotic stress response PROTEIN PHOSPHATASE 51 (OsPP51) transcript level was studied in CNP-treated seedlings compared to their controls. OsPP51 level was up-regulated to ~ 5-folds in the CNP-treated samples (Table S1) indicating improved abiotic stress compared to the controls.
Effect of different abiotic stresses in CNP-treated seedlings
Improved water relation parameters, bigger sized stomata and altered endogenous ABA content urged to study the effect of different abiotic stresses in CNP-treated seedlings. To this end, various stresses (i.e., cold at 18°C, high temperature at 35°C, salinity stress and induced drought using PEG) were imposed in separate experiments on seedlings with or without CNP in growth medium.
Changes in the RL under all these stresses were insignificant, hence, effects were studied in SL and RLN only. Seedlings showed 16.5% increase in SL and ~ 47.1% increase in RLN due to CNP treatment (Figure. 7A). This data served as the control for the abiotic stress treatments tested. Cold treatment for 7 d showed significant decrease in SL and RLN (Figure. 7A). While SL decrease ~ 64% without CNP, it decreased ~ 60% with CNP due to cold stress. RLN also showed higher decrease (~ 98%) without CNP and ~ 91.8% decrease with CNP due to cold temperature. These results indicated that CNP treatment could partly rescue the drastic effects on seedling growth during cold stress. PEG treatment creates osmotically induced drought environment at cellular level (Purbajanti et al., 2019). PEG treatment showed marginal insignificant increase in SL in both untreated and CNP-treated seedlings (Figure. 7A). Effect of PEG treatment showed significant contrasting response on RLN with or without CNP, i.e., 12.7% decrease without CNP and 9.2% increase with CNP treatment as compared with the control and CNP seedlings respectively. These results indicated that CNP treatment effectively counteracts the PEG induced cellular drought to show increase in RLN. Salt stress induced decrease in SL, RL and RLN both with and without CNP (data not shown). The % decrease due to salt stress was comparable, insignificant and within the range. High temperature treatment resulted decrease in SL, RL and increase in RLN (data not shown), however the differences were insignificant, therefore were not of interest. Despite germination, growth of seedlings was hampered under low temperature from 6th d-14th d. But, CNP-treated seedlings SL showed less effect as compared to the untreated ones (Figure.7B). These results indicated that CNP treatment has selective effects to countereffect the stress caused due to cold temperature or PEG treatment.
Countereffect of CNP-treatment on cold- and PEG-treatment induced stress in seedlings
To ascertain the seedlings growth effects observed under cold-stress and PEG treatment and the countereffects of CNP on it, various physiological and biochemical parameters were studied in 13 d-old (14 d-old in case of cold stress) seedlings with or without CNP (Fig. 7C-H). Cold stress resulted drastic decrease in chlorophyll content (~ 32.8-fold and ~ 18.2-fold in case of control and CNP-treated seedlings respectively), with higher amount of chl left in CNP-treated seedlings (Fig. 7C). Similarly, carotenoid content was also drastically reduced due to cold stress with ~ 16.1-fold and ~ 7.3-fold in case of control and CNP-treated seedlings respectively and lead to similar pattern of higher carotenoid content in CNP-treated seedlings left after cold stress (Fig. 7D). Relative water content (RWC) was not significantly different in the control seedlings with or without cold stress, however it was 7.1% increased (p ≤ 0.05) in the CNP-treated seedlings with cold stress when compared to without cold stress (Fig. 7E). Electrolyte leakage (EC) % was significantly higher in the control as well as the CNP-treated seedlings after cold stress (Fig. 7G) (1.78-fold and 2.1-fold respectively). Proline accumulation showed drastically different pattern among the CNP-treated and control seedlings (Fig. 7F). In the control seedlings, proline content was ~ 2.25-fold higher after cold stress as compared to the respective unstressed ones, whereas it showed nearly no change in their values in CNP-treated seedlings due to cold stress. Reactive oxygen species scavenging enzyme superoxide dismutase (SOD) activity was significantly higher in control (i.e., 5.7%), however in the CNP-treated seedlings SOD activity was not much increased after cold stress (Fig. 7H). These results indicated that the effect of cold stress was alleviated due to CNP treatment seedlings with higher chlorophyll, carotenoid, RWC, lesser SOD activity and nearly no more proline accumulation as compared to control seedlings.
PEG treatment for 6 d did not show significant decrease in chlorophyll in the control seedlings, whereas it showed ~ 34.1% increase (p ≤ 0.01) in chlorophyll in CNP-treated seedlings (Fig. 7C). Carotenoid content showed significant decrease by 27.7% in the control seedlings due to PEG treatment, whereas it showed nearly no change in the CNP-treated seedlings (Fig. 7D). RWC decreased marginally but significantly in both control and CNP-treated seedlings (6.8% and 3.8% respectively) after PEG treatment (Fig. 7E). Proline accumulation (Fig. 7F) and EC% (Fig. 7G) were insignificant in both control and CNP-treated seedlings after PEG treatment. Superoxide dismutase (SOD) enzyme activity was higher after PEG treatment in both control and CNP-treated seedlings, however the CNP-treated seedlings it was higher than the control ones (Fig. 7H). These results indicated that after PEG treatment, CNP-treated seedlings retained drought resistance characteristics but only in some parameters including chlorophyll, carotenoid content, RWC.
Assessment of cold and drought stress tolerance in rice plants in reproductive stage
To confirm the cold and PEG-induced-drought tolerance results obtained in the seedlings with CNP treatment, cold and drought stress was imposed in rice plants in the reproductive stage (after 4th CNP treatment).
Cold stress at 10°C for 10 d affected the cut leaves of untreated and CNP-treated leaves in a similar manner (Suppl Fig. 4). Yellowing of leaves, cold-induced lesions and drying progressed in similar fashion in untreated and CNP-treated leaves (Suppl Fig. 4A, 4B). Also, a recovery period of 24 h at room temperature was able to regain the greenness of leaves in untreated and CNP-treated leaves in a similar manner (Suppl Fig. 4C). However, chlorophyll and carotenoid content in the cold-treated leaves were higher in the CNP-treated leaves than the untreated ones (Suppl Fig. 4D-E). Further parameters were not analyzed as the leaves of untreated and CNP-treated leaves were apparently similar after cold stress.
Drought tolerance was tested by withholding water in control and CNP-treated pots (Fig. 8A). Water-stress (WS) for 36 h showed clearly resistant plant phenotype in the CNP-treated plants with greener stem, expanded and greener leaves than the control plants (Fig. 8B). After 48 h of water stress (Figure. 8C) and 24 h after re-watering, the CNP-treated plants had clearly more greener leaves (Fig. 8D). Chlorophyll content showed ~ 41% and ~ 61.2% decrease in both the untreated and CNP-treated plants respectively after WS. However, CNP-treated leaves retained the higher chlorophyll content after WS (Fig. 8E). Carotenoid content showed to be increased ~ 10 times after WS in both control and CNP-treated leaves with the later having the higher values after WS. RWC decreased drastically to nearly half of its starting value in the control plants, however it retained 82% in CNP-treated plants after WS (Fig. 8F). MDA content sharply increased by ~ 4.2-fold of its initial value in the control plants, however MDA content showed only ~ 2.2-fold increase in CNP-treated plants after WS (Fig. 8G), thus showing less MDA accumulation in CNP-treated plants than the control ones after WS. Proline accumulation showed opposite pattern in control or CNP-treated plants after stress. While proline content increased to ~ 1.5-fold in the control plants, it decreased non significantly in CNP-treated plants after WS (Fig. 8H).
Water relation and related parameters also investigated in the CNP-treated plants in comparison with control plants after WS. Leaf temperature increased by ~ 2.5°C in control while insignificantly in CNP-treated plants after WS (Fig. 8I). Relative humidity (RH) dropped ~ 37% in the control plants while it decreased insignificantly in CNP-treated plants after WS (Fig. 8J). Leaf CO2 concentration increased only ~ 15% in control plants, whereas in CNP-treated plants it increased to ~ 1.4 fold, maintaining the highest CO2 concentration after WS (Fig. 8K). Stomatal conductance was higher in CNP-treated plants in well-watered conditions. WS lead to drastic reduction of stomatal conductance in control plants up to ~ 20-fold. However, CNP-treated plants showed even higher (~ 53-fold) reduction of stomatal conductance under WS conditions (Fig. 8L). Transpiration rate presented similar scenario like stomatal conductance showing drastic reduction after WS (Fig. 8M). However, the CNP-treated and untreated plants had insignificant differences in their transpiration rate values both before and after WS. Vapor pressure deficit (VPD) was comparable between control and CNP-treated plants under well-watered conditions, however, WS increased the VPD to nearly two-fold with insignificant differences between control and CNP-treated plants after WS (Fig. 8N). These results indicated that CNP treatment in the rice plants could help to better withstand water stress by maintaining high chlorophyll carotenoid content, lower MDA, proline content, lower leaf temperature and lower stomatal conductance, transpiration rate.
Transcript expression analysis after water and cold stress in CNP-treated plants
To confirm the drought and cold stress tolerance phenotype equipped with biochemical and physiological characteristics in the CNP-treated plants during above stresses, transcript expressions of some ABA signaling, stomatal parameters and aquaporin genes were studied from flag leaf samples of CNP-treated plants after cold or water stress in comparison with their controls under well-watered condition (Fig. 8O-8T). Expression levels of ABA signaling components which are involved in stomatal closure such as OsSNRK1 and OsSNRK2 were down to ~ 0.36 and ~ 0.29-folds respectively after water stress in the CNP-treated samples as compared to their controls (Fig. 8O, 8P). ABA biosynthesis gene OsNCED1 expression was also down-regulated significantly by 10-fold in the CNP-treated samples (Fig. 8Q). Further, aquaporin gene OsPIP1-3 expression was found to be significantly up-regulated to ~ 3.9-folds in the CNP-treated samples as compared to their controls (Fig. 8R). These results indicated that CNP-treatment could alter ABA pathway, stomatal and water transport parameters to withstand water stress. To validate the cold stress tolerance parameters with CNP treatment observed at the seedlings stage and to access the cold tolerance at molecular level, transcript expression of INDUCER OF CBF EXPRESSION1 (OsICE1) (BHLH116) and DEHYDRATION RESPONSIVE ELEMENT BINDING PROTEIN 1D (OsDREB1D) were studied in rice flag leaf samples from CNP-treated and untreated plants after cold stress. OsICE1 expression was found to be significantly (p < 0.0005) down-regulated to nearly zero in the CNP-treated samples (Fig. 8S). OsDREB1D was also significantly (p < 0.01) down-regulated in the CNP-treated samples, however the down-regulation was 0.25-folds as that of the controls (Fig. 8T). These results confirmed that cold tolerances response genes were down-regulated in the CNP-treated leaf samples.
{kind=link}