CD45 magnetic separation followed by PIPseq captures the complexity of the immune microenvironment in the mouse brain
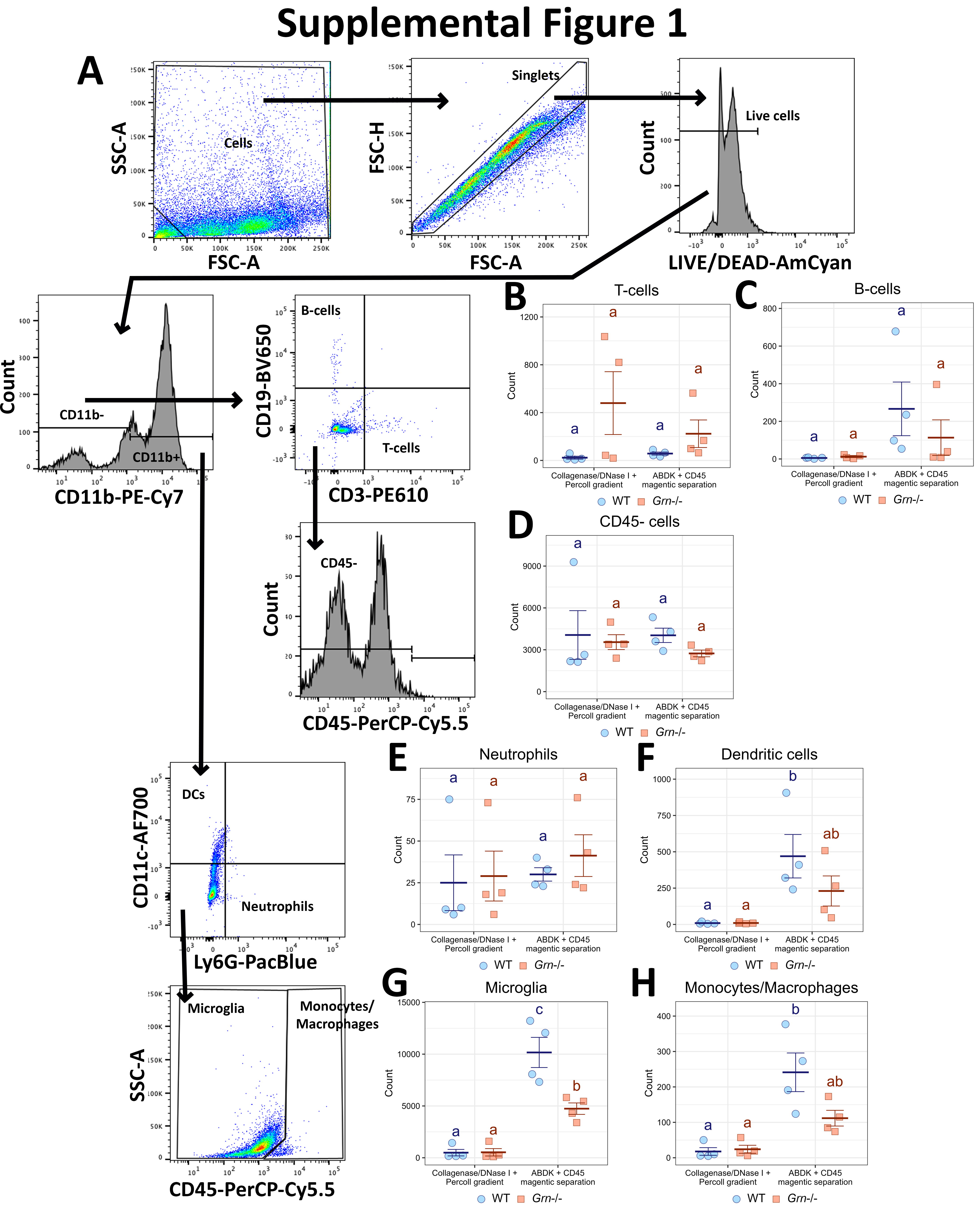
To interrogate central-peripheral immune crosstalk during neuroinflammation, we employed the well-established systemic endotoxin-induced neuroinflammation model by injecting C57BL/6J mice with LPS (1.5 x 107 EU/kg, i.p.) or an equivalent volume of sterile saline, and immune cells were isolated from brain tissue with ABDK and CD45 MS 24 hours post-injection. This brain dissociation strategy was selected over a strategy involving collagenase VIII digestion and Percoll-mediated selection of immune cells, which we and many others have used previously[13, 14], based on a preliminary experiment we performed that revealed higher yield and better purity of immune cells with the ABDK and CD45 MS (Figure S1). Since the use of enzymes to dissociate brain tissue has been shown to artificially induce immune signatures, mainly in microglia[41, 64], we included inhibitors of transcription, translation, and cell division or an equivalent volume of DMSO vehicle during dissociation as described previously[41] to examine the extent to which microglia and nearby immune cells are affected by this dissociation method. After isolation, 40,000 cells per sample were input to capture 20,000 cells in particle-templated instant partitions (PIPs), which were then lysed for downstream cDNA library preparation and sequencing[38] (Fig. 1A).
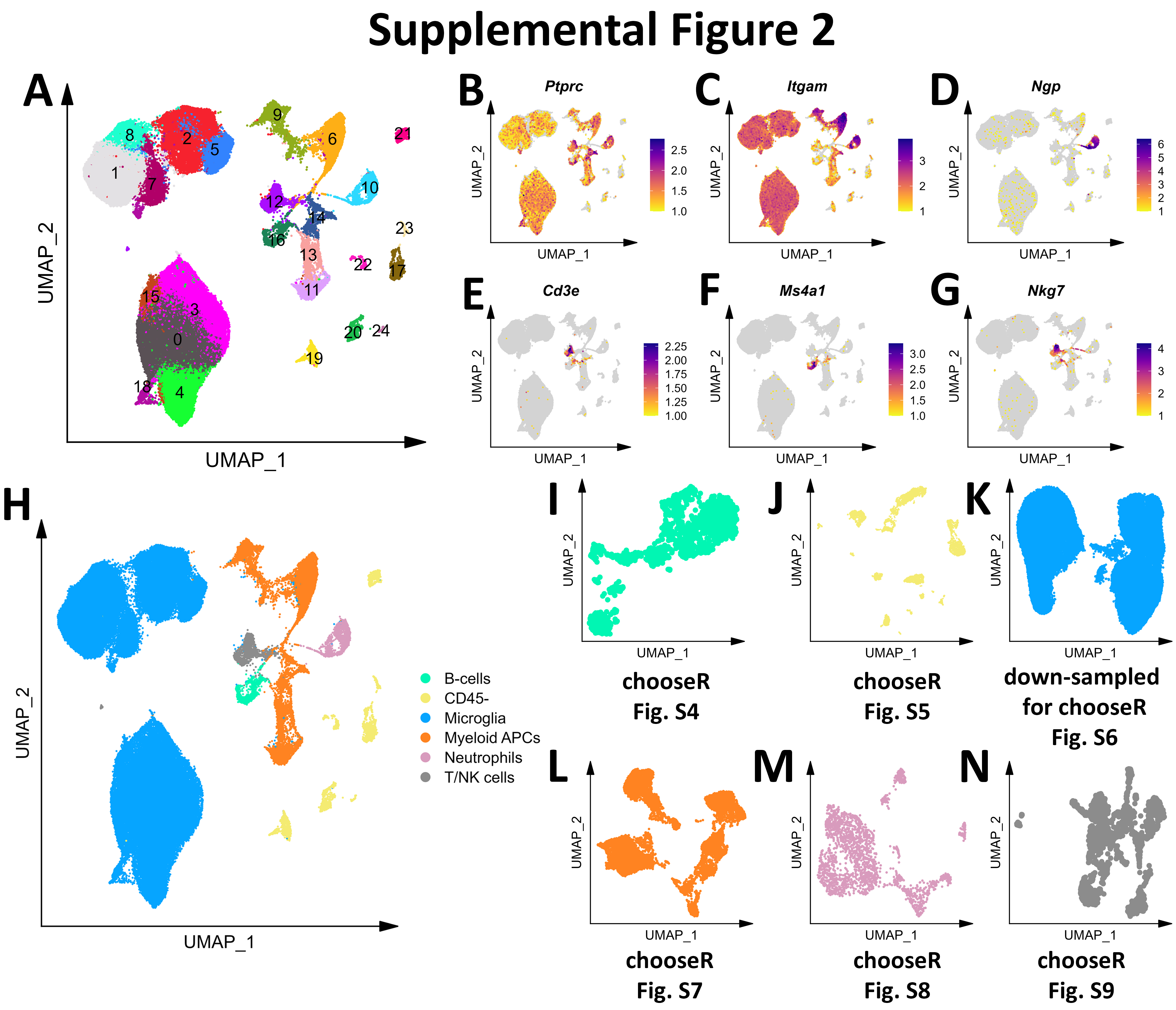
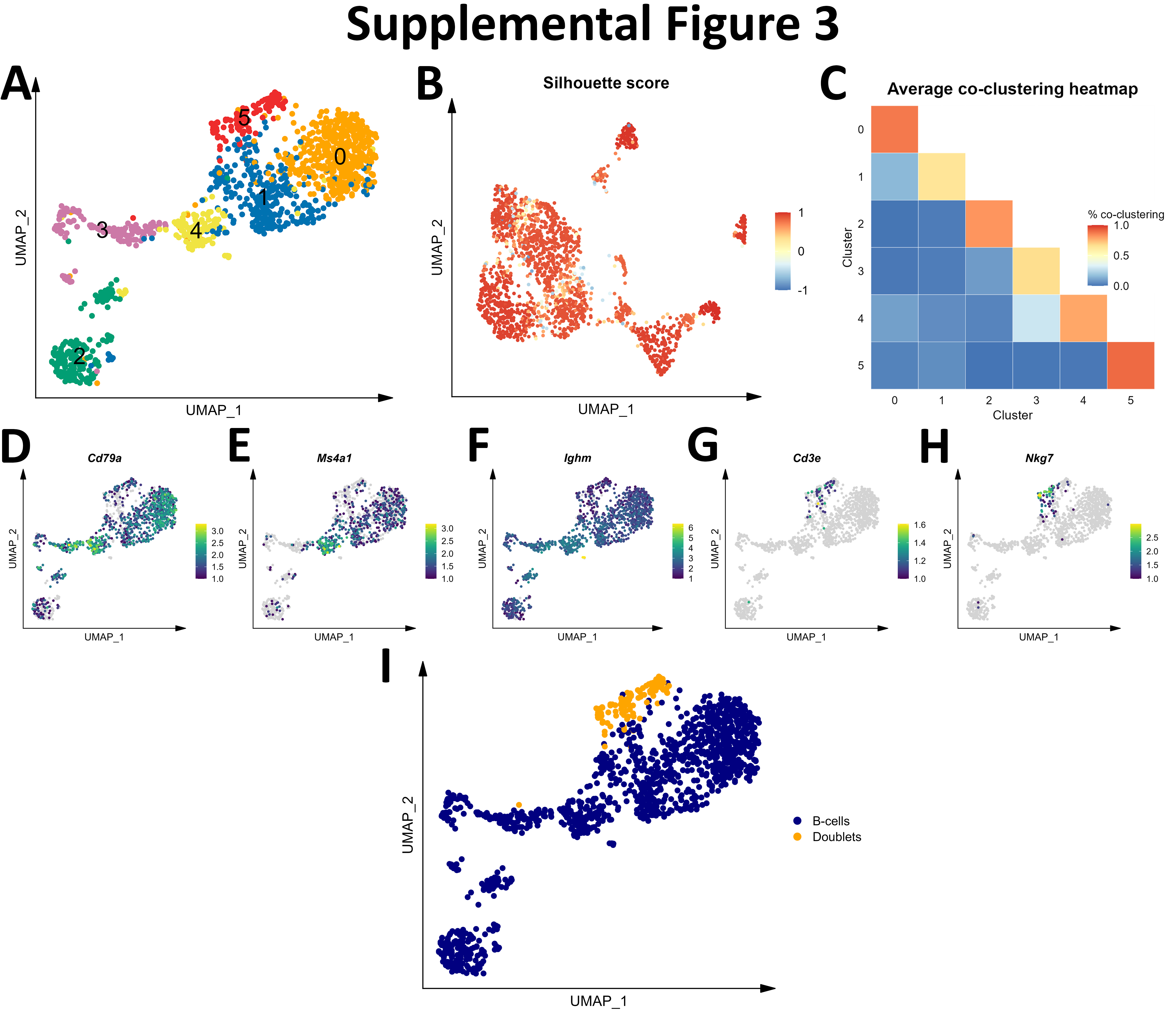
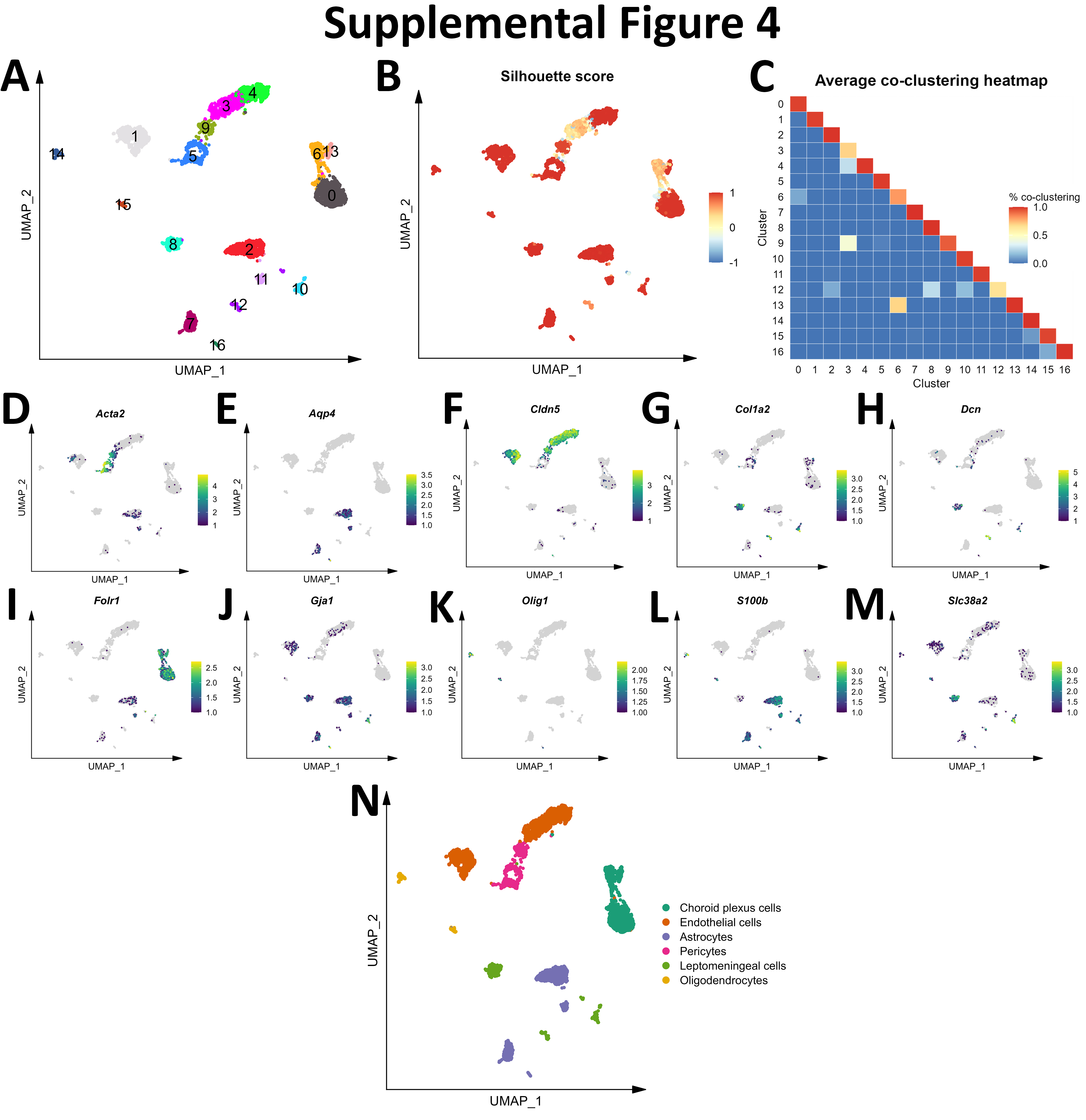
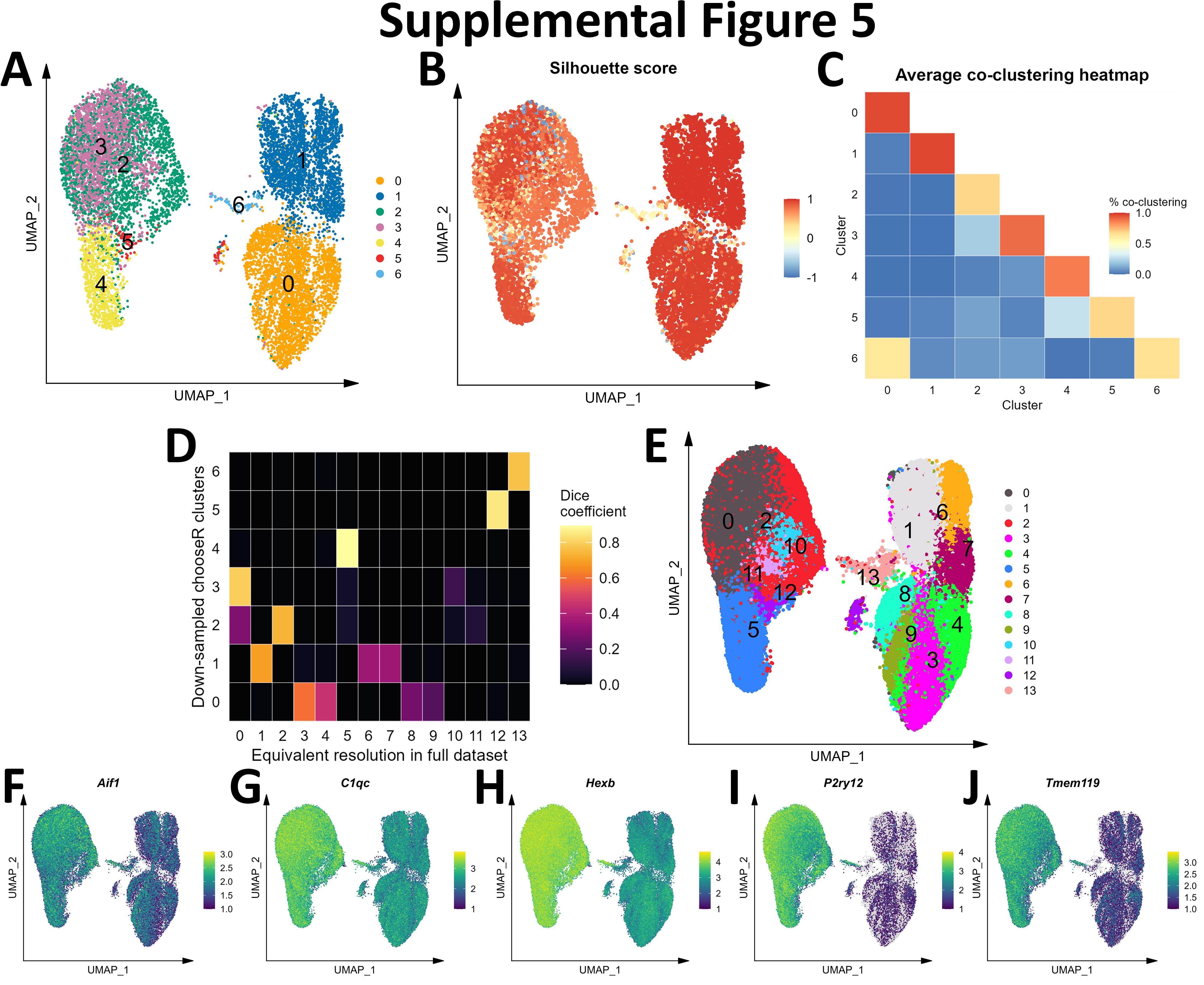
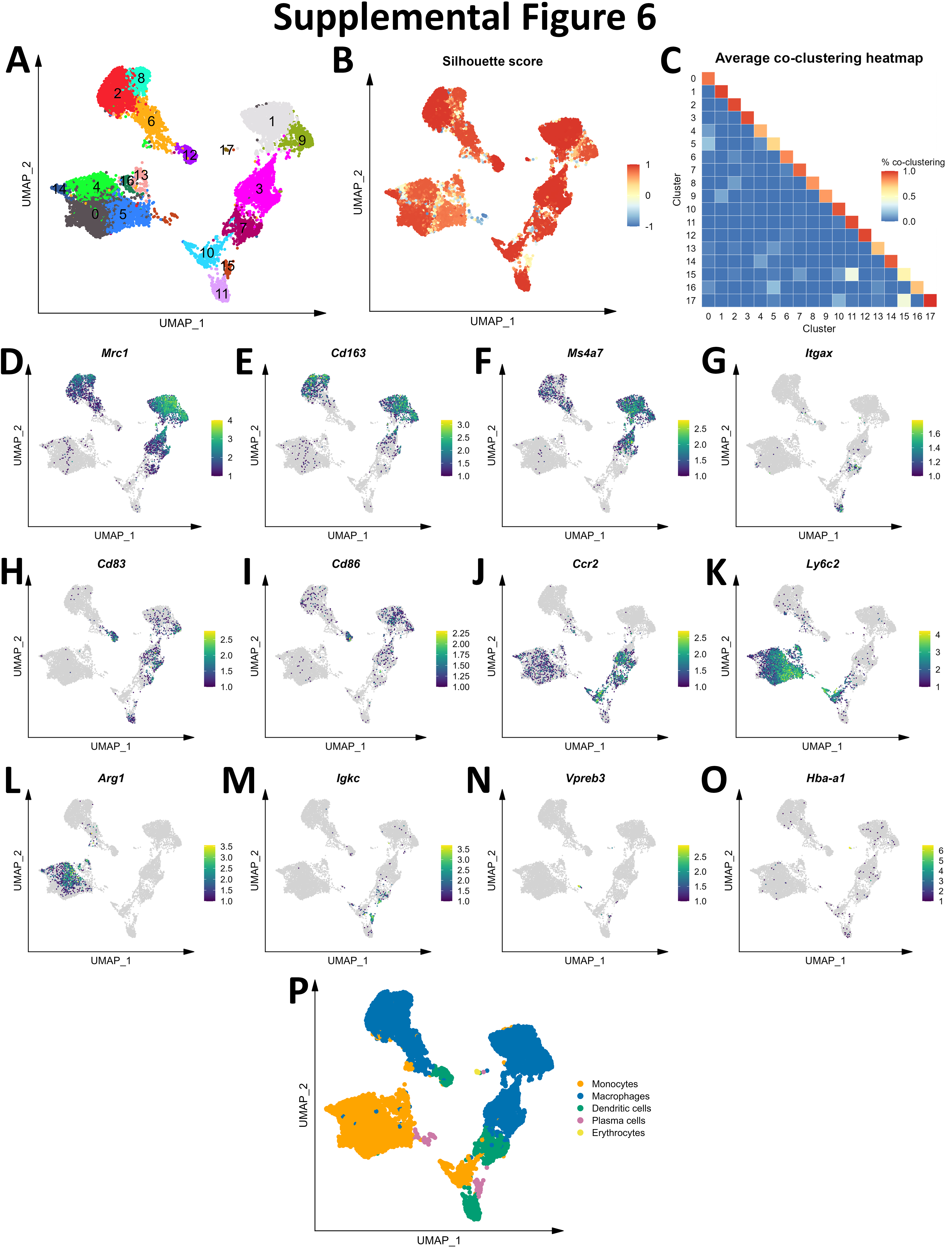
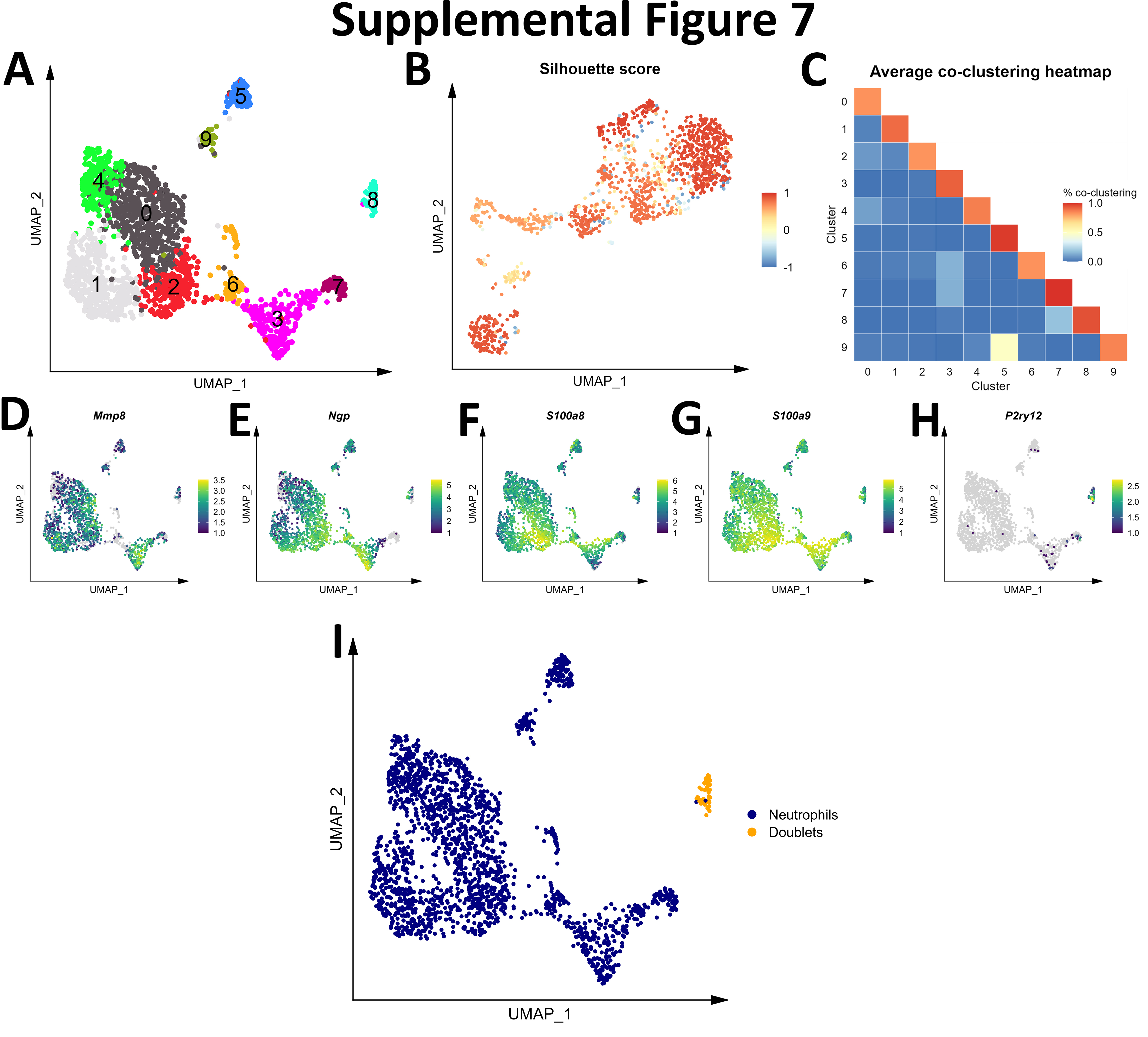
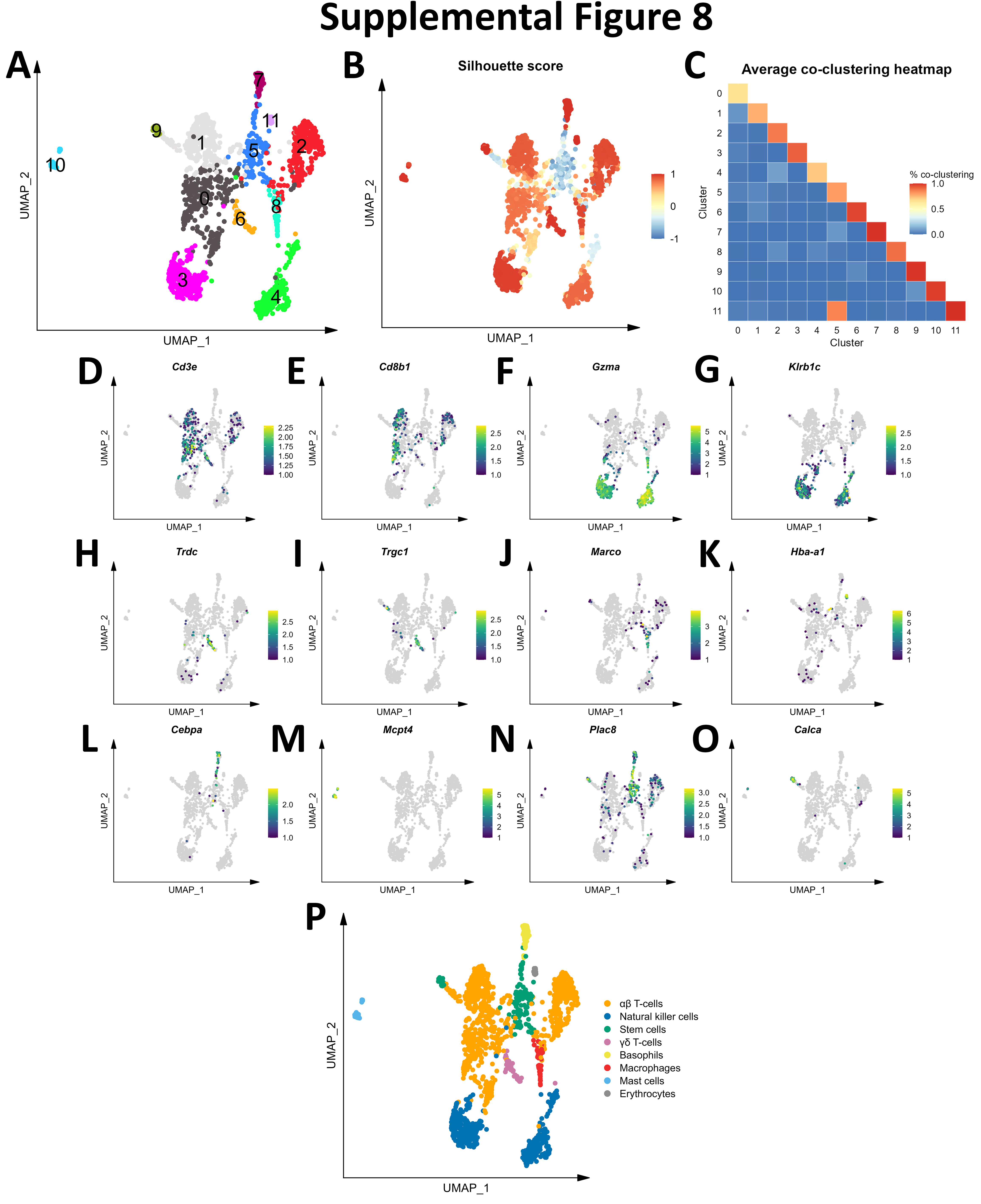
Initial graph-based clustering revealed 25 populations of cells, which were divided into six broad populations consisting of B-cells, CD45- cells, microglia, myeloid antigen presenting cells (APCs), neutrophils, and T- and natural killer cells (NKs), based on differential expression testing and the expression of canonical markers such as Ptprc, Itgam, Ngp, Cd3e, Ms4a1, and Nkg7 (Figure S2). We examined each cluster in detail with an iterative subsampling procedure and differential expression testing to confidently annotate cells (Figure S3 to Figure S8). The “B-cell” population was grouped into 6 subtypes (Figure S3) that each expressed B-cell markers such as Cd79a, Ms4a1, and Iglc3 (Figure S3D-F), although one cluster also expressed T-cell genes such as Cd3e and Nkg7 and was designated as a cluster of doublets and removed (Figure S3G-I). The CD45- population was grouped into 17 subtypes which expressed markers for astrocytes (e.g. Aqp4, Gja1, S100b), choroid plexus cells (e.g. Folr1), endothelial cells (e.g. Cldn5), leptomeningeal cells (e.g. Col1a2, Dcn, Slc38a2), oligodendrocytes (e.g. Olig1), and pericytes (e.g. Acta2) (Figure S4). The microglial population expressed classic microglial markers such as Aif1, C1qc, Hexb, P2ry12, and Tmem119 (Figure S5). The myeloid APCs were clustered into 18 subtypes (Figure S6A-C). Several cell types were identified based on marker gene expression, including macrophages (expressing Mrc1, Cd163, Ms4a7), monocytes (expressing Ccr2, Ly6c2, Arg1), dendritic cells (expressing Itgax, Cd83, Cd86), plasma cells (expressing Igkc, Vpreb3), and erythrocytes (expressing Hba-a1) were identified (Figure S6D-P). The neutrophil population consisted of 10 subtypes expressing neutrophil markers such as Mmp8, Ngp, S100a8, or S100a9 (Figure S7A-G). The microglial marker P2ry12 was expressed in one small cluster, which were classified as doublets and removed (Figure S7H-I). Lastly, the T- and NK cell population was grouped into11 subtypes which were annotated as αβ T-cells (expressing CD3 alleles, Cd8b1), NK cells (expressing Gzma and Klrb1c), γδ T-cells (expressing γ- and δ- T-cell receptor chains), erythrocytes (expressing Hba-a1), macrophages (expressing Marco), mast cells (expressing Mcpt4), basophils (expressing Plac8 and Cebpa), and stem cells (expressing many immature and proliferating cell markers (Figure S8).
In summary, we captured 20 different cell types from the mouse brain and its vessels that consisted of immune cells, including microglia, monocytes, macrophages, B- and T- cells, NKs, dendritic cells, and neutrophils, as well as a some non-immune and/or brain-resident cells (Fig. 1B). We also identified rare immune populations including basophils, mast cells, plasma cells, and γδ T-cells (Fig. 1C-F). Our cleaned dataset contained over 116,000 cells from only 8 mouse brains, over 90% of which were immune cell populations (Fig. 1G-H), demonstrating the efficacy of this CD45 MS-based approach in enriching the mouse brain and neurovasculature for immune cells.
Myeloid cells are vulnerable to an artificial activation state due to enzymatic dissociation
Enzymes to dissociate the mouse brain have been shown to induce artificial gene activation[41, 64]. Hence, we examined to what extent this already defined artificial microglia subpopulation was induced by the ABDK method and whether nearby circulating immune cells were affected. To determine whether the embeddings of cells differed based on inhibitor exposure, we performed a cross-entropy test[50] on immune cell subsets and the non-immune cell subset. This analysis revealed that granulocytes (composed of the neutrophil, basophil, and mast cell populations), macrophages, microglia, and monocytes were affected by enzymatic dissociation (Table 1). The low-dimensional embeddings of each of these populations in the inhibitor-treated and vehicle-treated samples are shown in Fig. 2A-D. The up-regulation of two published artifactual gene modules[41] can be seen in cells not treated with inhibitors during dissociation (Fig. 2E-L). To separate those artifactually activated cells, these three populations were clustered (Fig. 2M-P), artifact module expression was examined by cluster (Fig. 2Q-T), and the contribution of each experimental group to each cluster was calculated (Fig. 2U-X), revealing one cluster of granulocytes, three clusters of macrophages, three clusters of microglia, and one cluster of monocytes that were designated as artifactually activated (Fig. 2Y-BB). These clusters were removed ahead of biological interrogation of the LPS model.
Systemic administration of endotoxin activates central-peripheral immune crosstalk
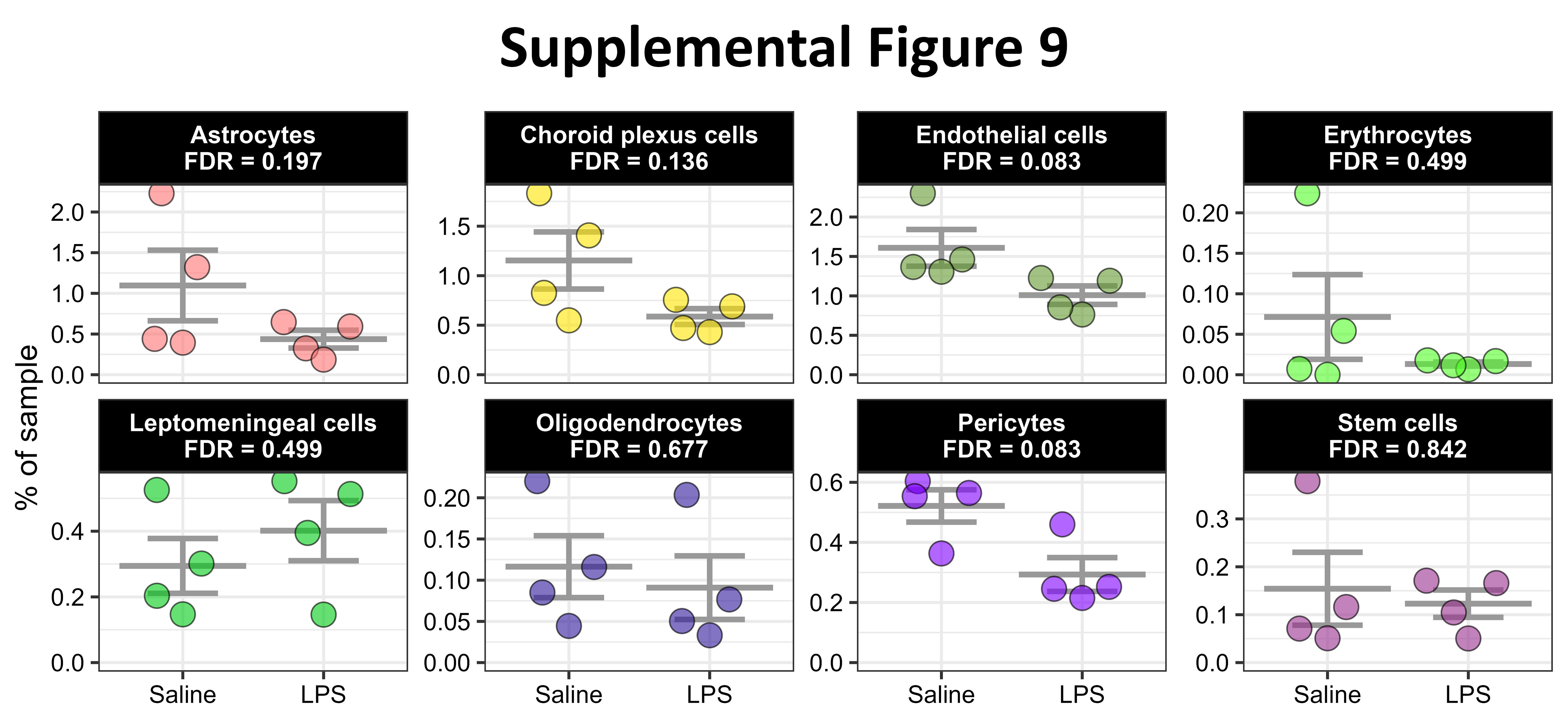
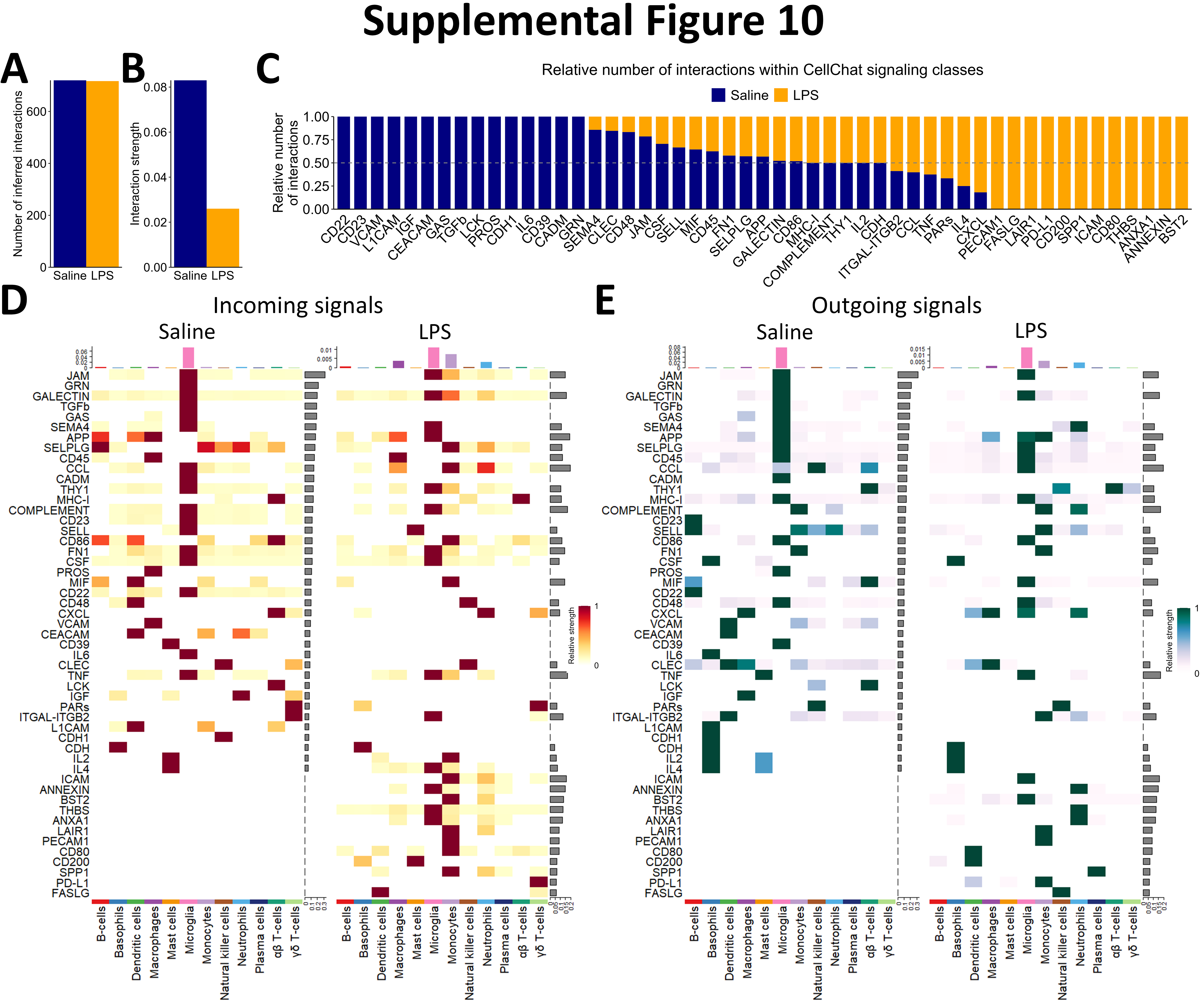
To begin dissecting the unique contributions of peripheral immune cells during LPS-induced neuroinflammation, we evaluated whether intraperitoneally administered LPS induces a change in the cellular composition of our brain immune-cell samples. We observed an increase in neutrophils and monocytes and a decrease in B-cells, dendritic cells, and plasma cells in/near the brain with LPS (Fig. 3A). The relative abundance of other immune cells and the non-immune cells captured here were not significantly affected by LPS (Fig. 3A, Figure S9). To determine the reason for these changes in immune cell composition with LPS, we inferred the communication of cells using multiple R packages, including CellChat[52] and MultiNicheNetR[53]. The overall strength of cellular communication was weakened by LPS (Figure S10A-B), and the nature of this cellular communication was modulated by LPS (Figure S10C), suggesting that circulating endotoxin alters immune cell communication between brain and blood. Indeed, the inferred communication between specific pairs of cells is modulated by endotoxin exposure (Fig. 3B-C). For example, macrophages were expressing fewer ligands with predicted receptors while neutrophils were expressing more ligands (Fig. 3C). Interestingly, microglia were expressing more ligands whose predicted receptors were expressed by peripheral immune cells, especially monocytes, NKs, and neutrophils, with LPS (Fig. 3C).
We hypothesized that microglia, the brain-resident cells in our dataset, would primarily govern the traffic of cells to and from the brain. We used MultiNicheNetR to predict the specific ligands expressed by microglia and the specific receptors expressed by those immune cells that are differentially associated with the brain after peripheral LPS exposure. This analysis revealed that chemoattractant and adhesion signals from microglia to B-cells and dendritic cells are largely down-regulated during peripheral LPS exposure. Specifically, several integrins, including Itgam, Itgav, and Itgb2 were downregulated in dendritic cells as were their putative binding partners Eng, F11r, and Cfh in microglia (Fig. 3D). Communication from microglia to dendritic cells and B-cells through transforming growth factor β (TGFβ) was disrupted by peripheral LPS exposure, and the adhesion of these cells to microglia through major histocompatibility complex class II (MHC-II) receptors (e.g., H2-DMa, H2-DMb1, and Cd74) and junctional adhesion molecules (JAMs; especially Jam2) was also decreased with systemic LPS exposure (Fig. 3D). Additionally, we inferred an elevation of several chemoattractant and adhesion signals from microglia to monocytes and neutrophils. The C-C chemokine, interleukin (IL)-1, and tumor necrosis factor (TNF) systems were inferred to be highly active between microglia and these two peripheral myeloid populations with peripheral LPS exposure, potentially creating a driving force for the migration of these cells. Additionally, the expression of adhesion molecules, especially Icam1 and Bst2, were upregulated in microglia as were their binding partners in monocytes and neutrophils (Fig. 3D). This analysis revealed novel potential binding partners that suggest an extravasation of mononuclear cells in the brain, including the interaction between tetherin (encoded by Bst2) and several paired-Ig-like receptors (Pira2, Pira12, and Pirb), orthologous to human leukocyte immunoglobulin-like receptors, and the interaction between heme binding protein 1 (Hebp1) and several formyl peptide receptors (Fpr2 and Fpr3). CellChat analysis corroborates the cell-specific modulation of signaling classes during peripheral LPS exposure. CellChat inferred a loss of incoming JAM and TGFβ and other adhesion molecules in dendritic cells and B-cells during peripheral LPS exposure, while monocytes and neutrophils were predicted to be receiving more interleukins, adhesion ligands, and chemokines (Figure S10D). In agreement with the above results, microglia were again predicted to be sending less TGFβ, more C-C chemokines, more TNF, and different adhesion factors (Figure S10E).
Finally, we examined the potential disruption of communication between microglial cells inferred by CellChat (Fig. 3C) using MultiNicheNetR. Of the top 30 differentially regulated ligand-receptor pairs, 26 were downregulated during peripheral LPS exposure, affirming that microglia-microglia crosstalk was disrupted by peripheral inflammation (Fig. 4A). Specifically, peripheral LPS exposure induced a loss of TGFβ signaling, integrin binding, and adhesion molecule expression between microglia. From the inferred loss of adhesion molecules, we hypothesized that LPS-exposed microglia would be more chemotactic. In support of this, a GSEA revealed that homophilic cell adhesion in microglia was disrupted by LPS and cell chemotaxis was strongly enriched in LPS-treated microglia (Fig. 4B-C). In addition, we hypothesized that in response to peripheral inflammation, microglia would be less homeostatic due to the overall downregulation of Tgfb1 and its receptors, as observed previously. A large gene set associated with tissue homeostasis was enriched in control microglia, in support of this hypothesis (Fig. 4B-C). Overall, peripheral LPS exposure disrupted the inferred signaling between microglia, antigen presenting cells, and other microglia. Instead, microglia expressed ligands for receptors highly expressed in peripheral myeloid cells to a greater extent, which may explain the greater frequency with which monocytes and neutrophils were found near the brain in our dataset.
Peripheral inflammation induces shared and cell-type specific transcriptomic programs in the brain
As the mobilization and communication of immune cells in the peripheral LPS exposure model is complex, we aimed to distinguish how different cell types contribute to peripheral LPS-induced neuroinflammation. We performed differential expression and gene set enrichment analyses in each cell type by creating pseudo-bulked datasets at the sample level and employing standard bulk RNA sequencing tools. Microglia displayed the greatest number of differentially expressed genes (DEGs), while adaptive immune cells and small granulocyte populations showed few DEGs, and peripheral phagocytes regulated a middling number of DEGs (Fig. 5A-B). We illustrate how many of these DEGs were unique to their respective cell type or shared by at least one other, showing a range of transcriptomic overlap depending on cell type (Fig. 5A-B).
To infer the functional state of each cell type, we employed the well-established gene set enrichment analysis (GSEA)[56, 57] on genes ranked by their differential expression due to peripheral LPS exposure. Several gene sets implicated in LPS signaling, cytokine response, protein synthesis, and metabolic activation were enriched in multiple cell types, suggesting that many immune populations, both in and near the brain, were metabolically and immunologically responsive to peripheral LPS exposure (Fig. 5C). However, gene sets associated with cytotoxicity, including the production of TNF, were enriched only in macrophages, monocytes, microglia, and neutrophils, while gene sets associated with cellular adhesion, cytoskeletal organization, and adaptive immune activation were disrupted in B-cells, T-cells, and NKs (Fig. 5C). Interestingly, neutrophils upregulated genes associated with phagocytosis and wound healing, while microglia and other peripheral phagocytes downregulated or showed no change in these pathways (Fig. 5C). These data suggest that the neuroimmune profile induced by peripheral LPS exposure is a product of the activity of many distinct immune cells, and certain inflammatory processes within this profile may be dominated by the activity of peripheral immune cells.
Finally, we curated lists of risk factors identified for Parkinson’s disease (PD) and Alzheimer’s disease (AD) in meta-genome wide association studies (meta-GWAS)[61–63] and examined the change in expression of these genes in each cell type due to an inflammatory stimulus. Microglia displayed regulation of many of these genes in response to peripheral LPS exposure, and some genes are uniquely regulated by microglia, including Gpnmb, Tmem163, Picalm, and Slc2a4 (Fig. 5D-E). However, many of these genes were also differentially regulated by peripheral myeloid cells in response to peripheral LPS exposure, including Bst1, Gch1, Lrrk2, Satb1, Apoe, Plcg2, Siglech, Sorl1, Spi1 (Fig. 5D-E). Most immune populations captured in this study up-regulated Ms4a6d and Tnip1 and downregulated H2-Eb1, Bin1, and Trem2 after peripheral LPS exposure (Fig. 5D-E). Interestingly, certain genes differed in the direction of their regulation between microglia and peripheral immune cells in response to peripheral LPS exposure, including Galc, Nek1, Abca7, and Cass4, while only peripheral populations regulated genes such as Camk2d, Ctsb, Grn, Map4k4, Abca1, Aph1b, Cd2ap, and Ptk2b. Overall, our findings reveal the existence of immune cell subset-specific expression of genes associated with risk for neurodegeneration in response to peripheral LPS, consistent with the growing perspective that genes that confer risk to neurodegenerative disease are linked to the peripheral immune system.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}