Multiple metabolic pathways are enriched in Breast Cancer tissue.
Gene expression analysis from breast cancer and non-cancerous breast tissue microarray data was curated from NCBI dataset GSE45827.(20) GSEA, using this microarray data and the MSigDB Hallmark gene set collection, uncovered multiple gene sets that are significantly enriched (FDR < 25%) in breast cancer samples. Of these, metabolic gene sets for glycolysis, and OXPHOS (Supplementary Tables 1 and 2) are significantly enriched (FDR < 25%) in breast cancer samples (Fig. 1A). The same gene sets also showed minor enrichment when comparing aggressive breast cancer subtypes (Luminal B, HER2+, TNBC) to Luminal A tumors (Fig. 1B). This enrichment analysis illustrates that gene sets for glycolysis (Fig. 1C) and OXPHOS (Fig. 1D) have significantly different expression in breast cancer and normal tissues. This preliminary analysis shows that glucose metabolism is altered in the cancerous state by varying degrees based on the cancer subtype.
miR-526b overexpression increases cell proliferation.
MCF7 and SKBR3 breast cancer cell lines were transfected to overexpress miR-526b. (12) Relative to MCF7-Parental, MCF7-miR526b maintains overexpression of pri-miR-526b by, on average, 20.7-fold. SKBR3-miR526b shows a 1306-fold increase in pri-miR-526b expression relative to SKBR3-Mock (Fig. 2A). These miRNA-high cells divide faster, and cell secretion turns media very acidic. Over 48 hours, MCF7-miR526b increased live cell number by 2.32-fold, while MCF7-Parental increased live cell number by only 1.40-fold over the same period. The same trend is seen in SKBR3, a cell line with a higher proliferation rate than MCF7. While SKBR3-MOCK increased cell number by 2.68-fold, SKBR3-miR526b increased live cell number by 3.99-fold over 48 hours (Fig. 2B).
miR-526b increases glucose metabolism.
Overexpression of miRNA in the transfected cell lines upregulates glucose metabolism as observed in MCF7 and SKBR3 breast cancer cell lines. One metric used to determine overall metabolic activity is ATP production, as ATP is the fuel for many reactions in the cell. MCF7-miR526b cells were observed to have a 1.81-fold increase in ATP concentration compared to MCF7-Parental cells. SKBR3-miR526b cells had a 2.07-fold increase in ATP concentration compared to SKBR3-MOCK (Fig. 2C). Thus, miR-526b overexpression moderately increases the ATP in cells.
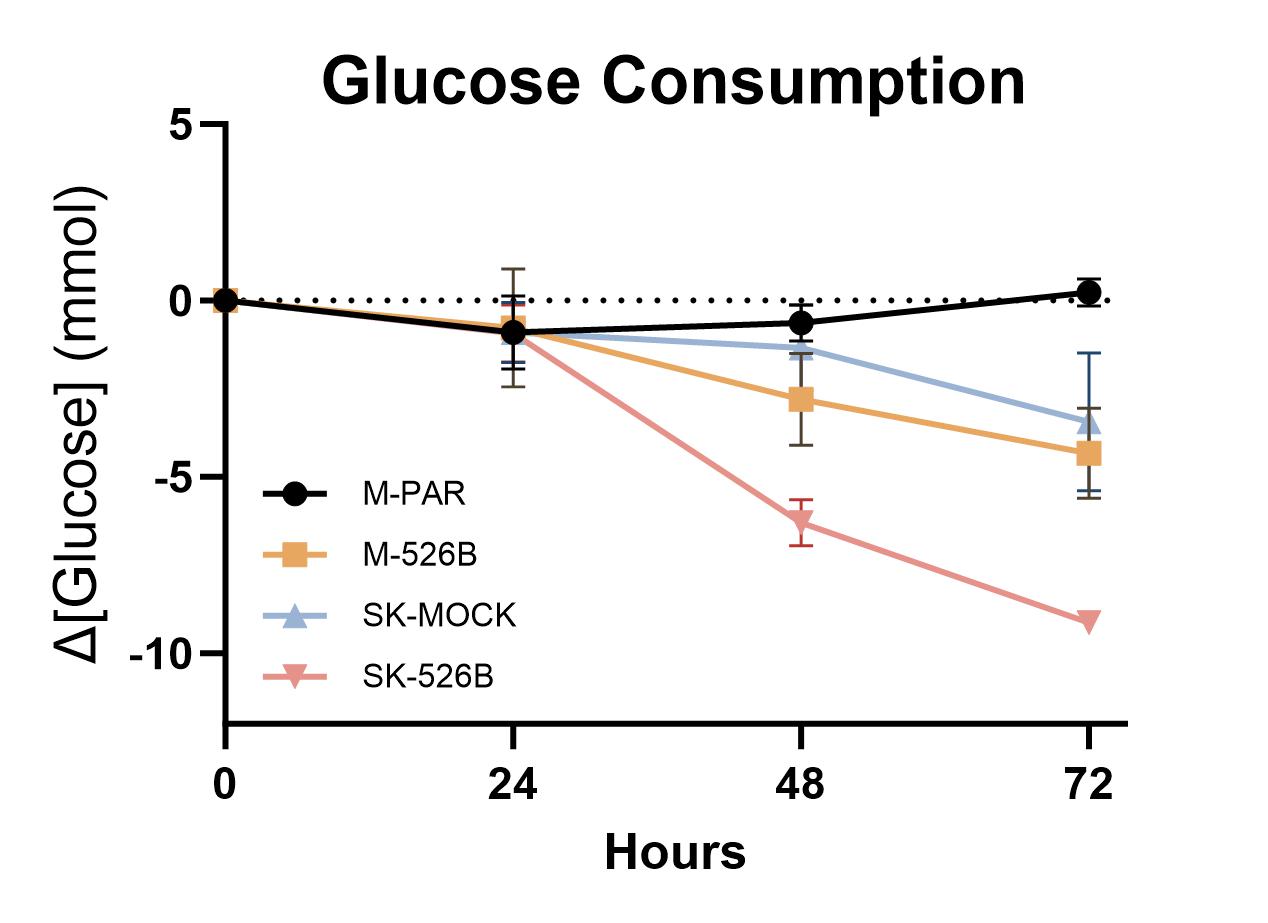
Next, glucose consumption was examined as glucose is one primary fuel for ATP synthesis. Glucose consumption was investigated by measuring the changes in glucose concentration in the cell culture media over time. For both MCF7 and SKBR3 cell lines, the miR-526b overexpressing line consumed significantly more glucose over 72 hours (Supplementary Fig. 1). Although we started with the same number of cells at t = 0hr, these cell lines have different proliferation rates; therefore, cell numbers must be considered over time. Glucose consumption per million cells was calculated to account for different proliferation rates. In general, the SKBR3 glucose consumption rate is higher than MCF7 cells. However, after considering differences in cell number, the overexpression of miR-526b did not increase glucose consumption significantly in either cell line (Fig. 2D). miR-526b expression resulted in more glucose consumed by cells overall, but this was shown to be a result of the increased proliferation rates of miRNA-high cells.
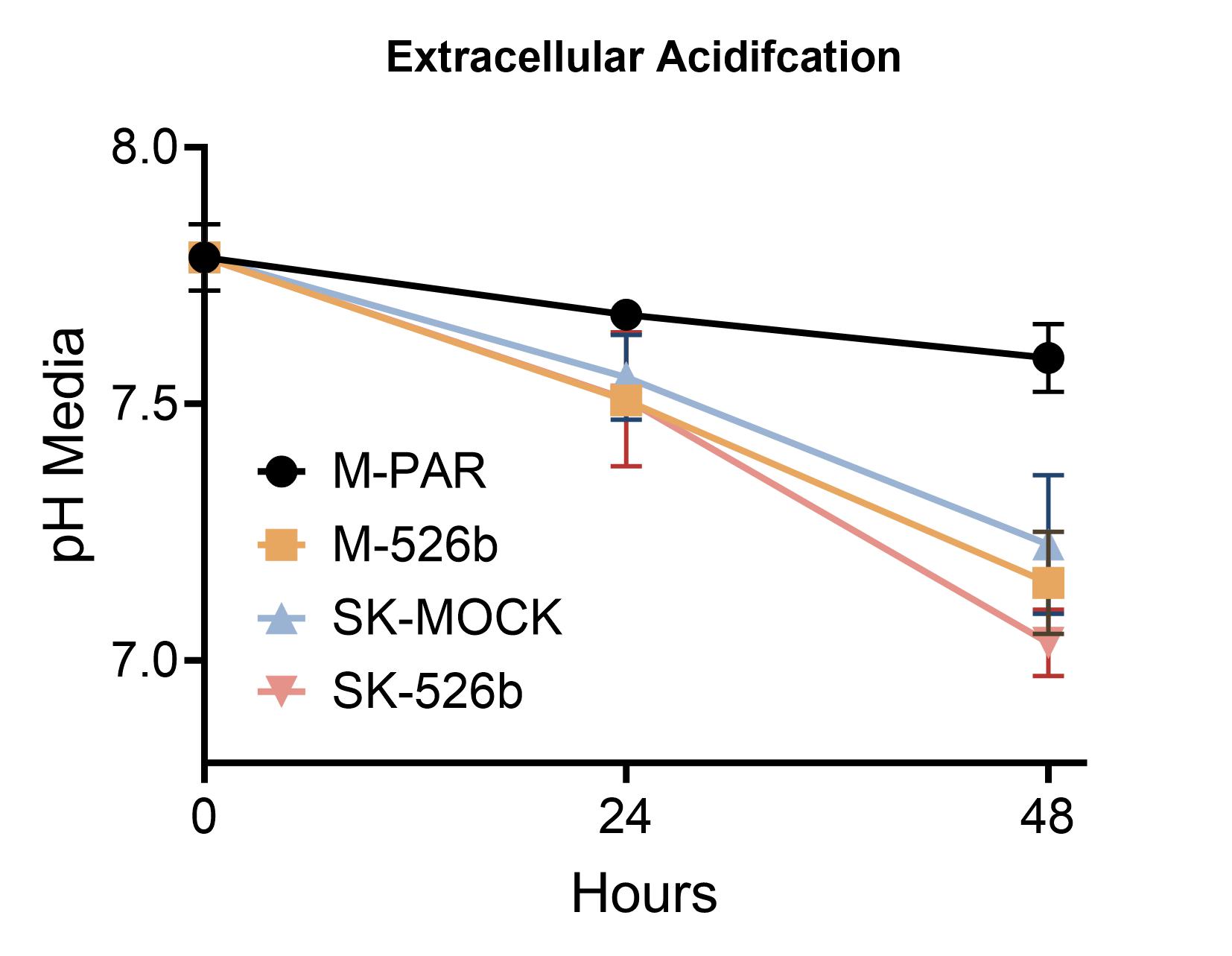
A decrease in extracellular pH often accompanies increased glycolytic metabolism. This is attributed to the increase in acidic byproducts of glucose metabolism, specifically lactate. We measured the culture media pH over 72 hours to see this effect. MCF7 showed the least changes in media pH (Δ = -0.2), but a significant reduction in pH was observed in MCF7-miR526b (Δ = -0.64). SKBR3-Mock cell media pH decreased by -0.56 over 48 hours, while SKBR3-miR526b showed a slightly more significant decrease (Δ = -0.76) (Supplementary Fig. 2). Because of the difference in cell proliferation rate, the increased acidification of the media could be due to cell division. To overcome this, a more sensitive method is used to monitor acidification in a reduced time frame. A fluorescent-based pH indicator is used, and over 2 hours, the extracellular acidification rate is calculated. In this case, the ECAR of MCF7-miR526b media is 0.42-fold less than MCF7-Parental. No significant differences were observed in the ECAR between SKBR3-MOCK and SKBR3-miR526b cells (Fig. 2E). miR-526b increases extracellular acidification; however, this is due to increased cell proliferation, not on an individual cell basis. When proliferation is normalized, miR-526b decreases ECAR in the MCF7 cell line and does not affect the SKBR3 cell line.
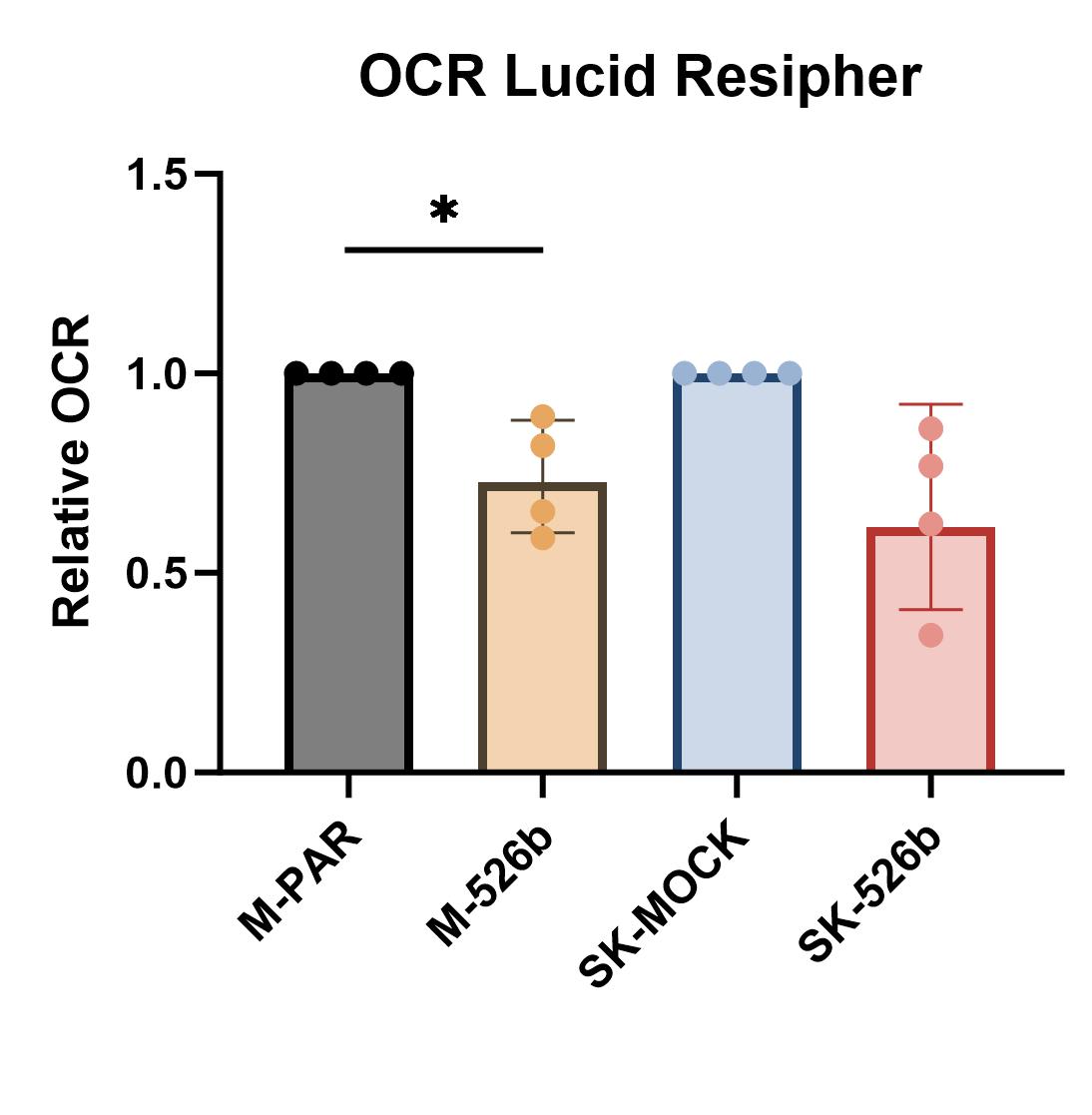
OXPHOS is an oxygen-dependent process; therefore, we can detect differences in OXPHOS between cell lines by measuring oxygen consumption. Increased oxygen consumption implies the cells utilize OXPHOS to synthesize ATP as O2 is consumed in the electron transport chain. For MCF7 and SKBR3, miR-526b overexpressing cells consumed less oxygen (Fig. 2F). The OCR was determined after interval measurements over 2 hours, limiting the effect of varying proliferation rates. A second OCR assay was performed over 48 hours. In this case, the real-time OCR at 48 hours was normalized to cell number by multiplying the initial cell number by the 48-hour proliferation rate identified in Fig. 2B. This long-term assay also shows that miR-526b overexpression leads to decreased oxygen consumption in both MCF7 and SKBR3 cells (Supplementary Fig. 3).
miR-526b dysregulates markers of aerobic glycolysis and glycolytic metabolism.
A panel of mRNA markers for glucose metabolism, Glucose transporter 1 (GLUT1), Hexokinase 2 (HK2), and Lactate dehydrogenase (LDHA) were screened to evaluate glycolytic and aerobic glycolytic pathways. Pyruvate dehydrogenase (PDHA1) and F1-F0 ATP Synthase subunit F1 (ATP5A1) were used as a marker for aerobic respiration. TP53-induced glycolysis and apoptosis regulator (TIGAR) were screened as potential targets of miR-526b that are involved in glycolytic regulation, as predicted by TargetScan V8.0 (Supplementary Table 3).(21, 22) An increase in expression of LDHA (2.31-fold) and PDHA1 (4.21-fold) was observed in MCF7-miR526b compared to MCF7-Parental, which naturally has a balanced metabolism (Fig. 3A). Other minor changes were observed in MCF7-miR526b, for example, HK2 (0.84-fold), ATP5A1 (1.31-fold), and TIGAR (1.45-fold). In contrast, SKBR3, which natively is more glycolytic than MCF7, shows notable differences. SKBR3-miR526b showed a decrease in glycolysis markers HK2 (0.38-fold) and GLUT1 (0.31-fold) compared to SKBR3-Mock. Also, marginal reductions in LDHA (0.79-fold) and ATP5A1(0.45) (Fig. 3B) were recorded. The differences observed in the two cell lines show how miR-526b can increase metabolic efficiency in diverse ways based on the cell's native metabolism.
Protein expression was measured for the predicted miR-526b target TIGAR and ATP synthase subunit ATP5A1 (full western blots in Supplementary Figs. 4–6). TIGAR protein was upregulated in MCF7-miR526b and SKBR3-miR526b (Fig. 3C-D). ATP5A1 protein expression was unchanged in MCF7-miR526b and significantly downregulated in SKBR3-miR526b. This indicates that miR-526b might be involved in aerobic glucose metabolism and energy production in breast cancer.
The effect of glycolysis inhibition on MCF7, SKBR3, and miR-526b overexpressed cell lines.
The sensitivity to specific metabolic inhibitors is examined to better understand the changes in metabolic profile. By targeting or inhibiting glycolysis using 2-deoxy-d-glucose (2-DG), we can evaluate metabolic plasticity or the cell’s ability to adapt to continue to meet high energy demands and survive. To examine the impact of this inhibition, relative ATP concentration, cell viability, and proliferation are measured. When exposed to 2-DG treatment, SKBR3-MOCK cells showed a more significant decrease in ATP than MCF7-Parental. This is expected as SKBR3 is more glycolytic; therefore, the inhibition of glycolysis would impact ATP yield. Both MCF7-miR526b and SKBR3-miR526b were less affected by 2-DG treatment than their miR-low counterparts (Fig. 4A), indicating miR-526b might help cells switch between glucose metabolism pathways.
Cell viability is the proportion of live cells following treatment. After glycolysis inhibition by 2-DG, all cell lines had high viability, although overexpression of miR-526b increased viability marginally in both MCF7 and SKBR3 cell lines (Fig. 4B). Viability illustrates cell death, yet the differences in total live cells after treatment stood out. To quantify how the drugs are affecting cell proliferation, the number of live cells following treatment is compared to the number of live cells following a vehicle control treatment over the same time. This better illustrates the inhibitors' effect on cell growth and division rate when the treatment is not causing cell death. For both cell lines, the 2-DG treatment has less impact on live cell count in the miR-526b overexpressed lines. Therefore, with miR562b overexpression, the cells were less affected by 2-DG treatment and better-maintained proliferation (Fig. 4C). The fluorescent images (Fig. 4D-G) show both viability and proliferation. In the miRNA-overexpressed cell lines, there are more live cells (blue) and fewer dead cells (red). This indicates that miRNA-overexpressed cells can more effectively overcome glycolysis inhibition.
The effect of OXPHOS inhibition on MCF7, SKBR3, and miR-526b overexpressing cell lines.
OXPHOS is inhibited using Oligomycin, which blocks mitochondrial ATP synthesis. To examine the impact of this inhibition, relative ATP concentration and cell viability/proliferation is measured. When exposed to Oligomycin treatment, all cell lines showed a slight increase in ATP production, with no significant differences identified between cell lines or miR-526b status. An increase in ATP is expected as the cancer cells can adapt and produce ATP, often in higher yields under hypoxic conditions. The inhibition of OXPHOS mirrors hypoxic conditions in that both prevent cell respiration and the electron transport chain. ATP was decreased with the combination treatment, where cells were exposed to glycolysis and OXPHOS inhibitors. Cell lines with high miR-526b maintained ATP production better in this case. (Fig. 5A).
After OXPHOS inhibition by oligomycin, overall cell viability remained high. MCF7-miR526b showed slightly better viability (97.3%, 91.4%) than MCF7-Parental (88.6%, 85.6%). The opposite was true for SKBR3. SKBR3-miR526b showed decreased viability (96.5%, 89.7%) compared to SK-MOCK (98.6%, 93.5%) (Fig. 5B). Also, MCF7 showed the same result regarding cell proliferation. MCF7-miR526b cells showed higher proliferation (0.83, 0.72) than MCF7-Parental (0.72, 0.59). SKBR3 cells showed the same result; SKBR3-miR526 b cells (0.92, 0.81) showed higher proliferation in response to oligomycin than SKBR3-MOCK cells (0.77, 0.73) (Fig. 5C). This indicates the miR-high cells better maintain their growth after OXPHOS inhibition, but this is less sustainable in SKBR3-miR526b cells as there is a higher proportion of cell death.
The fluorescent images (Fig. 5D-G) show both viability and proliferation. In the MCF7 miRNA-overexpressed cell lines, there are more live cells (blue) and fewer red cells (dead). For SKBR3, there are more live (blue) cells and dead (red) cells in the miR-high treated conditions. So, miR-526b overexpression in MCF7 and SKBR3 cell lines enhances the metabolic plasticity of breast cancer cells by maintaining cell proliferation.
COX2, EP4, and miR-526b contribute to metabolic dysregulation
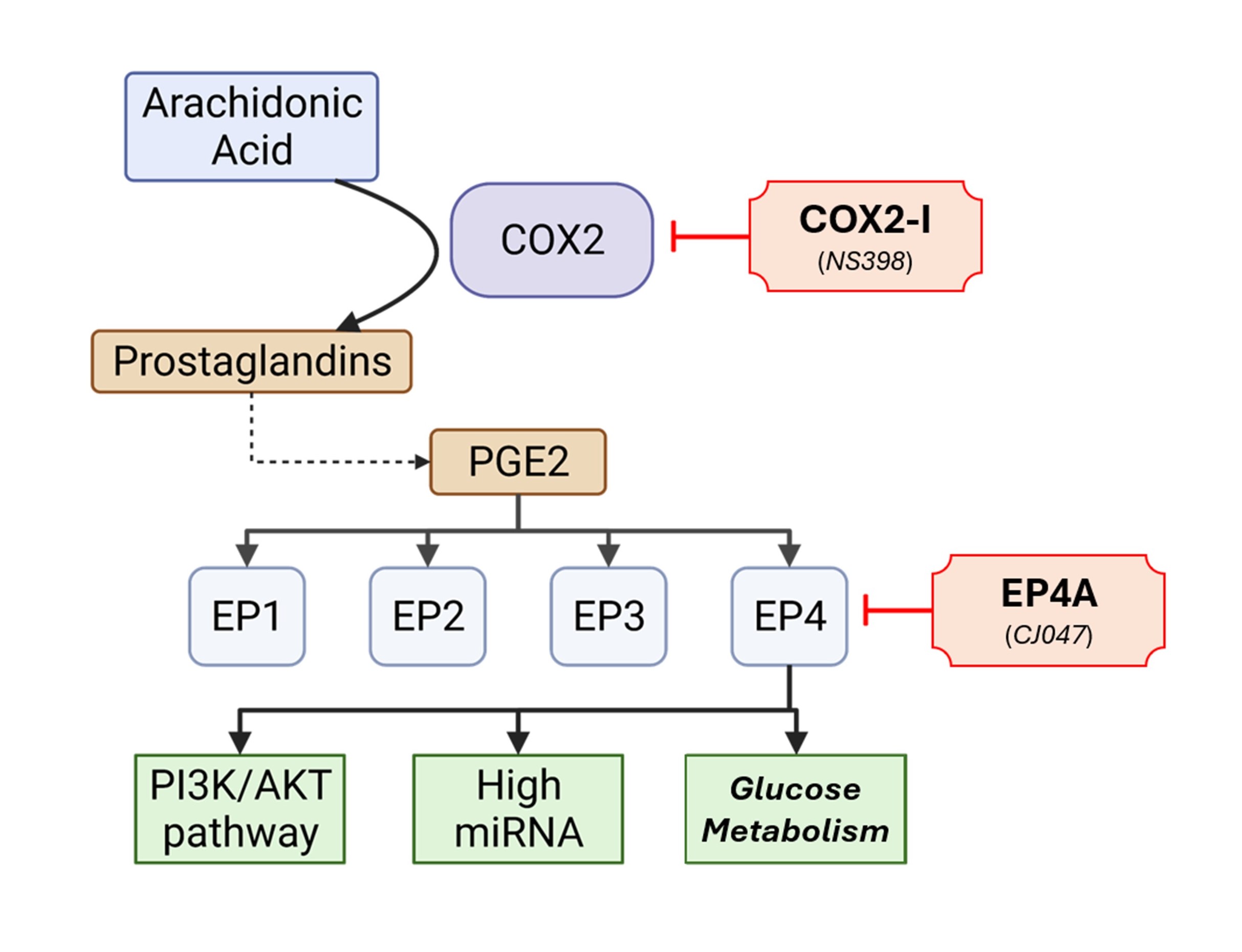
We discovered that miR-526b was upregulated in Cyclooxygenase-2 (COX2) overexpressing breast cancer cells, and many oncogenic functions of miR-526b could be abolished with a COX-2 inhibitor. COX2 is part of an inflammatory pathway that results in the production of Prostaglandin E2 (PGE2), which binds to G-protein-coupled receptors (GPCR) EP1, EP2, EP3, and EP4, each having different cellular localizations and downstream signal transductions.(23, 24) COX-2 upregulates miR-526b via the EP4 signaling pathway, and miR526 b-induced functions can be disrupted with the use of NS398, a COX-2-specific inhibitor, and CJ047, an EP4-specific antagonist (Supplementary Fig. 7).(10) COX-2 is the master regulator of miR-526b.
MCF7-COX2 cell line shows upregulation of miR-526b with an average increase in pri-miR-526b of 32.0-fold compared to MCF7-Parental (Fig. 6A). MCF7-COX2 shows an increased proliferation rate like that observed with miR-526b overexpression. (25) MCF7-COX2 cell number increased by 2.87-fold over 48 hours, while MCF7-Parental only increased cell number by 1.4-fold (Fig. 6B). MCF7-COX2 showed transcriptomic changes similar to MCF7-miR526b regarding RNA expression of metabolic markers. Compared to MCF7-Parental, MCF7-COX2 showed marginal downregulations in glycolytic markers HK2 (0.75-fold), GLUT1 (0.84-fold) and LDHA (0.76-fold). Genes associated with OXPHOS were upregulated: PDHA1 (2.93-fold) and ATP5A1 (14.0-fold). TIGAR gene expression was also marginally upregulated in MCF7-COX2 cells (1.59-fold) (Fig. 6C). TIGAR and ATP5A1 protein expression was significantly upregulated in MCF7-COX2 cells compared to MCF7 (Supplementary Fig. 6). TIGAR showed 2.45-fold upregulation, and ATP5A1 showed 3.37-fold upregulation compared to MCF7-Parental cell line expression (Fig. 6D, E).
miR-526b expression and function can be inhibited with COX2 inhibitor and EP4 antagonist treatment.
Following the norm of miR-526b function, 48-hour treatment with COX2 inhibitors and EP4 antagonist reduces the expression of pri-miR-526b by 0.64-fold and 0.36-fold, respectively, in MCF7-COX2 cells (Fig. 6F). Similarly, this treatment also decreases cell proliferation. The relative proliferation rate in COX2-i treated cells is 0.74 relative to vehicle control treatment. MCF7-COX2 cells treated with EP4A show a proliferation rate of 0.93 relative to vehicle treatment (Fig. 6G).
Interestingly, the observed miR-526b regulation of metabolic markers can be reversed with COX2 inhibitor and EP4 antagonist treatment. LDHA, PDHA1, and ATP5A1 gene expression are reduced in MCF7-COX2 cells following treatment with COX2-i and EP4A (Fig. 6H). Therefore, treatment with a COX2 inhibitor and EP4 antagonist inhibits miR-526b expression and reverses miRNA-induced glucose metabolism.
{kind=link}
{kind=link}
{kind=link}
{kind=link}