2.1 Study Area
We conducted data collection and analysis at three scales, 1) the larger rice field landscape, 2) the local rice field complex, and 3) the interior of rice paddies (Figure 1). The larger rice field landscape is a rectangular 4200km2 area centered around the rice fields but which includes the Doñana National and Natural parks, Veta la Palma aquaculture ponds, and adjacent agricultural areas within the Guadalquivir river delta (Figure 1); this area encompassed >90% of all LBBG GPS fixes during the study period, and was used as the study area for all static habitat selection analysis. The rice field complex is a zone of intensive rice cultivation to the Northeast of Doñana National Park, and is the largest rice production area in Spain. It lies within a Biosphere Reserve (Green et al. 2018) and comprises several hundred small (4-35 ha) polygonal paddies divided by dikes, small roads, and irrigation canals and portions of the Guadalquivir river. The rice field complex includes all landcover falling within a 250m buffer of rice paddy edges. It also includes larger urban settlements, fragments of shrubland and forest, relict natural marshland and extensive areas of dryland agriculture including cotton fields and fruit orchards. Finally, we examined habitat features in the interior of individual paddies, which we defined as the area at least 45m interior of the paddy edge (Toral et al., 2011). We used this analytical scale for our dynamic habitat analysis of space use with respect to within-paddy condition.
Habitat availability in rice paddies follows a consistent pattern according to the annual rice harvesting cycle, which we divided into three phases. During the pre-harvest stage, all paddies contain unharvested rice. The active harvest phase begins with rice harvest starting in late September and early October, and paddies are tilled or plowed after harvest, and finally flooded. Flooded conditions in rice paddies persist from October to December or January, depending on ambient precipitation and evaporation rates (Toral et al., 2011). The active harvest phase is marked by high heterogeneity in habitat types, because the harvest, tilling, and flooding occur in a spatiotemporally staggered and piecemeal fashion because of limited harvesting equipment, leading to asynchronous transitions between within-paddy conditions (Toral et al., 2011). The post-harvest phase begins in mid- to late-January, when all fields have typically been harvested; for the purposes of this study, the post-harvest phase of the overwintering season ends with the departure of overwintering LBBG in mid-March.
2.2 GPS Tracking Data
We used data on the movements of LBBGs tracked using Global Positioning System (GPS) trackers (University of Amsterdam Bird Tracking System, UvA-BiTS, Bouten et al., 2013) from breeding colonies in north-west Europe. The GPS trackers measured spatial position (mean position error of 3m for 60s intervals, 30m for 600 second intervals) and acceleration (tri-axial accelerometer; Bouten et al., 2013). Birds were captured and fitted with trackers under license on their breeding grounds in Northern Europe (United Kingdom, Thaxter et al. 2019; Belgium, Stienen et al., 2016; the Netherlands, Shamoun-Baranes et al., 2017). The UvA-BiTS data are stored in a centralized database (http://www.uva-bits.nl; Bouten et al. 2013). Prior to analyses, all GPS data were checked for erroneous velocity measurements (>80km/hr, the upper limit observed for this species; Shamoun-Baranes et al., 2017), potentially unreliable fixes (with fewer than 4 satellites used for geopositioning), and bursts (periods of sampling where a large number of fixes were taken over a short period of time), to avoid biasing estimates of space use. Erroneous or unreliable data points were excluded. We examined the fix interval (duration between fixes) for all GPS trackers in the database to investigate temporal structure in sampling, and sub-sampled the data to a common interval (20 minutes) to avoid bias in habitat selection calculations (Shamoun-Baranes et al., 2011); this filtering step retained ~80% of fixes (Additional File 1). We estimated the Haversine distance (spherical distance between geographical coordinates of GPS fixes) between each consecutive point, and examined the mean and total daily distance traveled for all gulls throughout the overwintering period for seasonal patterns. Finally, we calculated trajectory speed as distance/duration.
We selected data for the 2016-2017 winter season because that year had a relatively high number of tracked birds appearing in the study area (n=18), more low cloud-cover satellite images available than other years, and precipitation levels close to the average values of the past 20 years. We used all GPS data for the period of August 15, 2016 to March 15 2017 (hereafter overwintering period) within our study area, omitting all individuals with less than 1000 GPS fixes prior to data filtering and subsampling. These individuals were removed because they either had a small number of days (<14) present in the rice field landscape during the study period or in one case had sampling intervals that were temporally coarse (>3 hr) yielding poor information on fine-scale habitat use and movement patterns. In total, excluded individuals accounted for <4% of all fixes recorded in the rice field landscape during the 2016-2017 overwintering season, while the remaining dataset contained >45,000 GPS fixes from 6 individual gulls (five individuals tagged in Zeebrugge, Belgium and one on Skokholm island, U.K.).
We created two data subsets based on GPS fixes that coincided with the dates of available imagery for our dynamic habitat analysis. The first of these consisted only of data collected on the same day as each available remotely sensed image (24-hour window), and consisted of 3,082 fixes, and the second (expanded dataset) included fixes from one day before and one day after each image (72-hour window), for a total of 9,185 fixes; this is slightly less than three times as many as the 24-hour window dataset, because two images were two days apart from the proceeding image, leading to an overlap in their 72-hour windows. The 24-hour window dataset has a lower probability that harvest status changed between image collection and a given GPS fix within that window but has reduced statistical power due to a smaller sample size, while the 72-hour window dataset increases sample size and statistical power but at slightly increases the risk of including GPS fixes collected under different harvest conditions.
For each of the 6 remaining birds, we also used the UvA-BiTS virtual lab (Bouten, 2018) online client to project and visually check trajectories of unfiltered data for each gull prior to quantitative analysis. This was done by subsampling data points to 3-hour intervals and then visually examining GPS trajectories for coarse-scale movement behavior within the rice field landscape (e.g., movements between the rice paddies and other landscape features, or repeated daily movements to features outside of the rice field landscape). Any forays outside of the rice field landscape (Fig. 1) were visually examined at a 20-minute interval resolution to identify additional destinations during the overwintering season.
2.3 Remote-sensing Imagery, Classification, and Ancillary Spatial Data
We downloaded and classified a total of 14 images (ten from Landsat 5, 7 and 8 and four from Sentinel-1), with dates ranging from 2 September 2016 to 17 February 2017 (days 7 to 175) to detect and quantify the dynamic availability of habitat resources for gulls at the rice paddy scale throughout the overwintering period (hereafter “dynamic habitat analysis”). All images were geometrically and radiometrically corrected according to Aragonés et al. (2005). We classified harvest status landcover types within each paddy in the rice field complex for each available image date. Following Toral et al. (2011), we restricted the classification of rice paddies to the interior of each paddy by excluding a 45m buffer inwards from their outer edge to avoid spectral confusion with land-cover categories that are not found within paddies (e.g. shrubland, roads). Notably, this means that use of features outside of paddy interiors (e.g., dikes and roads between paddies) were quantified only in our static habitat analysis.
We classified these clipped images using K-means unsupervised classifications with the RStoolbox package (Leutner et al., 2019) in R 3.5.2 (R Core team, 2019). We chose a value of K based on the largest consensus among a suite of 30 indices using the Nbclust package (Charrad et al., 2015) in R. Where no clear consensus among metrics was available, the parsimonious (smallest K) competing value was chosen. We used the resulting image with the highest number of habitat classes present as the basis for thematic classes used in all other classified images. We attempted at minimum to classify harvest status conditions that were relevant to habitat use by gulls (see also Martín Vélez et al. 2020), specifically green vegetation or unharvested rice (low-density foraging activity), recently harvested rice (large amounts of active foraging), tilled rice (active foraging), and flooded areas (loafing, sleeping, and occasional foraging). These classes were assigned using knowledge from field visits and reference images.
We detected 7 thematic classes for harvest status within rice paddies: bare ground, dry tilled, wet tilled, deep flooded, shallow flooded, harvested rice, and green vegetation (either pre-harvest rice or other plants colonizing fields post-harvest). Deep-flooded and shallow-flooded paddies were detected based on spectral signatures, wherein deep-flooded fields showed a signature strongly indicative of water, while shallow-flooded paddies showed some mixture of water and bare ground, indicating that land was reflecting some light through a shallow layer of water; water depth in neither case likely exceeds 30cm, so the terms ‘deep’ and ‘shallow’ are based only on relative differences in water depth.
In addition to image-based classifications of harvest status, we generated an additional map of habitat resources that were unlikely to change throughout the wintering season for an analysis of static habitat features at the rice field landscape scale. For this we used a reclassified version of landcover data from the SIPNA database (Sistema de Información sobre el Patrimonio Natural de Andalucía, Junta de Andalucia, REDIAM, 2019), which included rivers and dikes, roads, non-rice agriculture, urban areas, natural and artificial ponds, and other more permanent features in the larger rice field landscape. We simplified thematic classifications for this dataset to match behaviorally-relevant habitat types for LBBG for a final habitat map of the larger rice field landscape (for extent, see Fig. 1). This involved combining classes that presumably had no impact on gull biology into single classes, for example, combining industrial and residential development into “urban”, and different types of fruit orchards into “non-rice agriculture”. Areas known to be rice paddies were lumped into a single class, “Unclassified Rice” to reflect birds using rice paddy interiors in our static habitat analysis (for days without imagery and subsequent harvest status information; Additional File 2). Finally, we generated a shapefile consisting of a 250m buffer around all rice paddies, to be used for determining whether or not given GPS fixes were occurring within the rice field complex or in another part of the rice field landscape (e.g., landfills, more distant sections of the Guadalquivir river, aquaculture ponds or natural marshes, Fig. 1).
2.4 Accelerometer Data
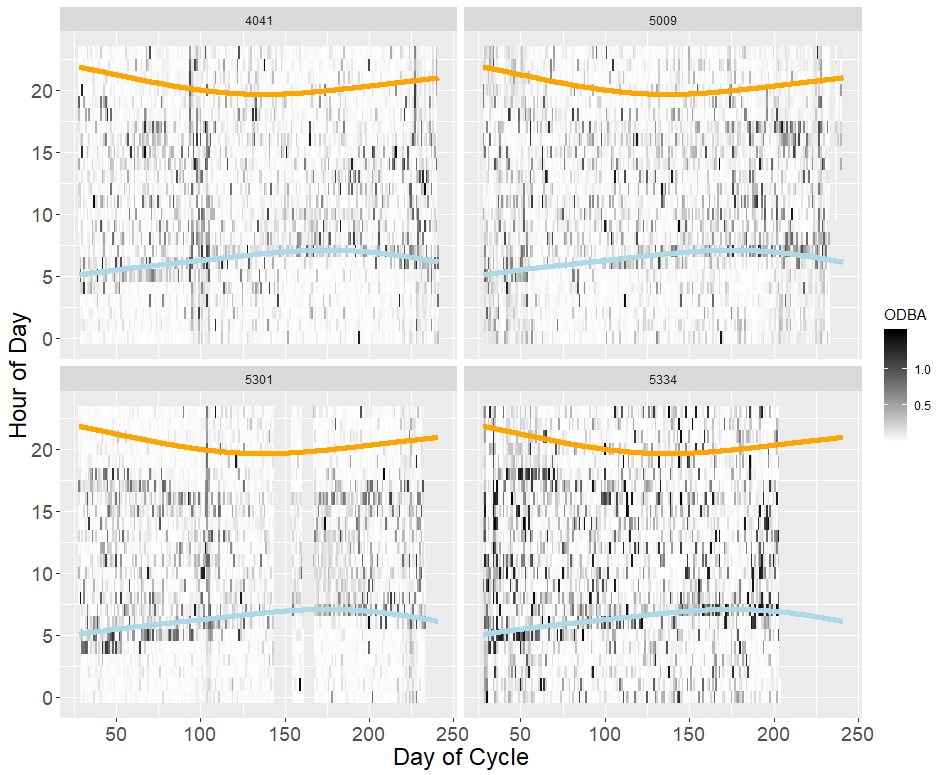
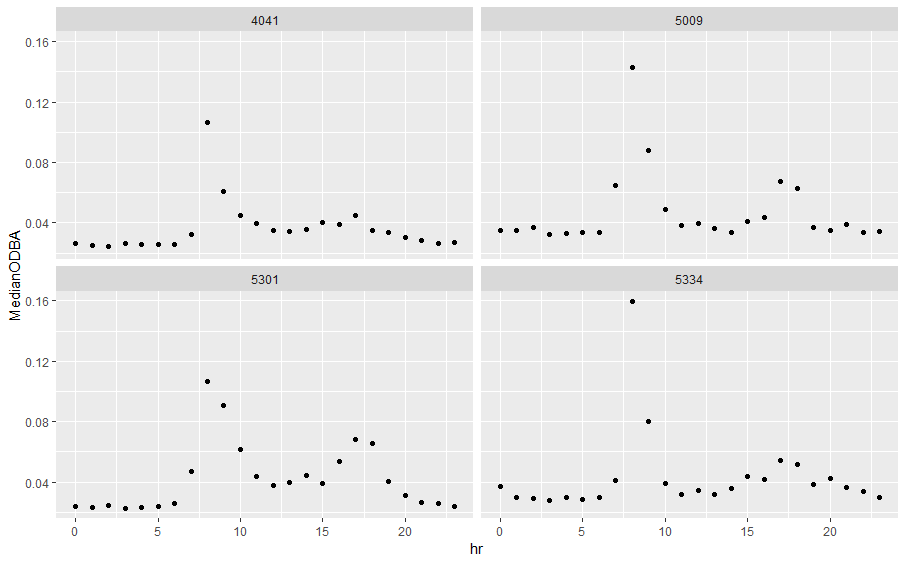
Tri-axial accelerometer data (1-second segments of acceleration data in three dimensions collected concomitantly with GPS fixes at 20hz frequency) were available for four of the tagged gulls. We removed accelerometer segments with more or less than 1-second of data to avoid bias, yielding a dataset of 10,641 accelerometer segments. We converted all accelerometer measurements to G’s (equivalent to gravity at the Earth’s surface, 9.8m/s2), and calculated mean and standard error along each directional axis for each segment in order to calculate dynamic acceleration. Finally, we calculated overall dynamic body acceleration (ODBA) as the sum of all dynamic accelerations for each segment (Wilson et al., 2006; Nathan et al., 2012). We classified accelerometer data using the random forest model and behavioral classes for LBBG generated by Shamoun-Baranes et al. (2016). These classes were terloco (terrestrial locomotion, walking), flap, exflap (rapid, active flapping), soar (gliding flight), manouvre (flapping and soaring combined), peck, , sit/stand (resting or inactive with the body level), walk, float, and boat (resting on a moving object). We associated exflap, peck, and terloco with feeding and food-searching related behaviors, flap, manouvre, and soar with travel, and boat, float, and sit/stand with resting. These accelerometry-based behavioral classifications differ from those outlined for field observations (below), which were based only on visual identification and observation. We validated accelerometer classifications by checking the correspondence of classified behaviors with other spatial characteristics of the associated GPS fix for a subset of 100 fixes (e.g., whether floating was associated with points in water, or whether instantaneous velocity was high enough for points classified as soaring or flying).
2.5 Field Observation Data
We conducted behavioral observations of overwintering LBBG in the rice field complex to complement and validate the GPS and accelerometer data. We collected observational data between 0900 and 1800 hours from early November to the end of January during the 2018-2019 overwintering period (days 75-157). We constructed an ethogram of seven behaviors that were readily distinguishable in the field: walking, searching-foraging, flying, swimming, bathing, sitting, and standing. We classified all observations according to these mutually exclusive behavior classes. Behavioral data consisted of instantaneous scans as well as focal bird observations. We performed instantaneous scans on paddies that contained LBBG and scanned the entire paddy (including surrounding dikes and airspace above it) for a 1-minute interval and counted the number of gulls performing a certain behavior. To reduce the potential for temporal autocorrelation in behaviors, repeated instantaneous scans in the same site were conducted no less than 10 minutes apart. For observations of focal birds, a single individual was monitored for up to 30 minutes, recording the amount of time spent on each behavior. Observations were ended if line of sight of the bird was lost. We also recorded time of day, harvest status (unharvested, actively being tilled, or harvested, and not flooded, partially flooded, or flooded), weather, and GPS location of the approximate paddy centroid. We conducted 142 instantaneous scans and 22 focal follows (totaling approximately 10h of observation) across 70 locations and 7 days of observation.
2.6 Space Use Analyses
We classified all GPS points according to whether they fell inside or outside of the rice field complex, and whether they occurred during the day or night (according to daily sunrise and sunset times, using the R package RchivalTag (Bauer, 2018). We additionally classified all points by land use based on our static habitat map (rice field landscape scale) using a simple overlap point extraction (R package ‘raster’; Hijmans & van Etten, 2012). For our dynamic habitat analysis, we classified all fixes that fell within the interior of rice paddies by harvest status using the same method. We conducted this analysis for both the 24-hour and 72-hour window data subsets. Accordingly, all GPS fixes received a static habitat classification, and our imagery-based subsets (24- and 72-hour windows) received specific information on harvest status at the paddy scale for our dynamic habitat analysis. We converted all date-time stamps of GPS fixes to a “day after arrival”, where day 1 was the day of the first bird’s arrival into the rice field landscape (for the 2016/2017 overwintering season, this was 27 August, such that 1 January 2017 was day 128). We present all results in both calendar date and day after arrival.
We conducted space use analyses at population and individual scales, using simple overlays and point-selection functions. The relatively small position error for fixes filtered for quality (~3m) lends credence to this type of analysis, although such errors could result in biases against detecting space use in thin, linear features like smaller roads and dikes. We conducted separate analyses for GPS fixes during the day and night to examine differences in gull behavior across the daily cycle. We also conducted analyses on data that were subset according to the three phases of the overwintering harvest cycle: pre-harvest (days 1-50, or August to mid-October), active harvest (days 50-140, or mid-October to mid-January) and post-harvest (days 140-202, or mid-January to mid-March, when the last bird left the rice field landscape), based on Toral et al. (2011), our own remote sensing datasets, and field observations across years.
We used the static habitat (rice field landscape scale) map to compare the selection of rice field habitats vs. non-rice field habitats and investigate the role of non-paddy habitat features embedded in the rice field complex, while we used the image-based data subsets for a dynamic analysis of how rice paddy harvest status influenced the use of the rice paddy interiors by LBBG. For both analyses, we calculated Manly selection ratios for habitat types following Fletcher and Fortin (2018) and using the widesII function in the package AdehabitatHS (Calenge and Basille, 2019). We calculated availability by randomly sampling 10,000 points from our habitat layers using the sampleRandom function in package Raster for classified remotely-sensed images (Hijmans et al., 2019) and the over function in package sp for our vector-based static habitat dataset (Pebesma et al., 2018). Availability and selection were calculated on a by-image (thus 24-hour or 72-hour) basis for the dynamic (image-based) habitat dataset. We calculated separate selection ratios by phase of the overwintering season and day versus night, as well as their factorial combination. Manly selectivity ratios were considered significant when their 95% confidence intervals did not overlap zero. Because results for dynamic habitat selection analyses were not qualitatively different between our 24- and 72-hour window data subsets, all results for habitat selection among paddies of different harvest status are presented using the 72-hour window subset (~10,000 GPS fixes).
2.7 Abundance Data
We assessed the abundance of LBBG in the rice field complex using two independent time series of LBBG abundance. First, we used aerial survey data collected monthly during 2016-17 by the monitoring team of the Doñana Biological Station (EBD-CSIC), following the methods explained in Rendón et al. (2008). Briefly, counts of the Doñana wetlands are performed by a single trained observer during a ~3 h flight in a small plane along a ~450km planned route at 40-250m elevation, including about 60% of the rice field complex on the west side of the Guadalquivir river (Mañez et al., 2012). We included only count data from August to March. From the air, Yellow-legged gull (Larus michahellis) and LBBG cannot accurately be separated, but the numbers of Yellow-legged gulls in rice fields are comparatively trivial during winter (CBvR & AJG, pers. obs), averaging <10% of all counted gulls during winter. . Yellow-legged gulls make up <1% of all gulls counted in river surveys within the rice field complex (see below), making aerial counts a reliable index of LBBG abundance.
We also analyzed data from weekly daytime (0800 to 1700 hours) counts of gulls that were made via line transects on a boat along the Guadalquivir river from Spring 2008 to Fall 2009 (the only period for which these survey data were available). We selected only those survey observations collected within the rice field complex and calculated total daily abundances for each survey date during the 2008-2009 overwintering period.
2.8 Analyses of Behavior
We analyzed the prevalence of different behaviors within the rice fields and throughout the overwintering season using classified accelerometer data (in 2016-17), protocols of focal birds (in 2018-2019) and instantaneous scans in the field. We used chi-squared tests to detect significant relationships between paddy-scale harvest status and behavior prevalence and then used correspondence analysis (using the packages FactoMineR; Husson et al., 2019) to visualize these relationships in low-dimensional space. For behavioral data collected in the field, behavior prevalence was measured by all gulls engaging in a certain behavior (focal birds), or the number of gulls performing that behavior at given point in time (instantaneous scan). For accelerometer data, behavioral prevalence was the proportion of fixes (among all fixes that had classified accelerometer data, controlled for sampling interval) classified as that behavior. We assessed the potential for autocorrelative bias in behavior by repeating calculations with instantaneous scan data including and excluding replicates from the same day and site. For accelerometer data, we conducted analyses separately using our static habitat dataset (landscape-scale, all data points) and our dynamic habitat dataset based on available imagery (paddy-scale, 24- and 72-hour subsets).
{kind=link}
{kind=link}