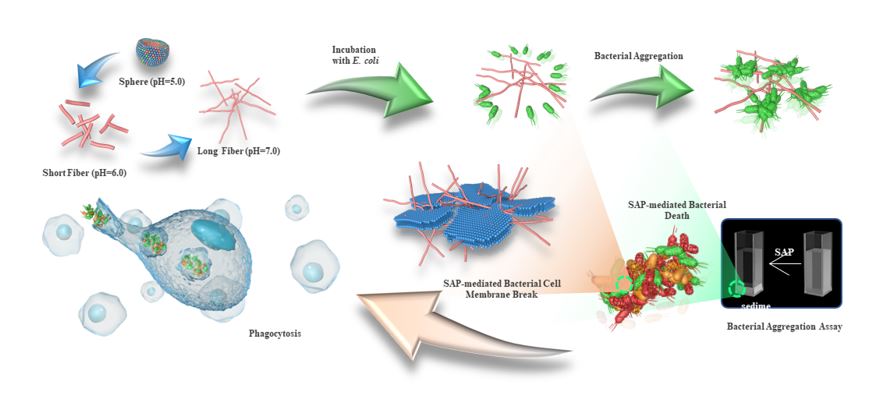
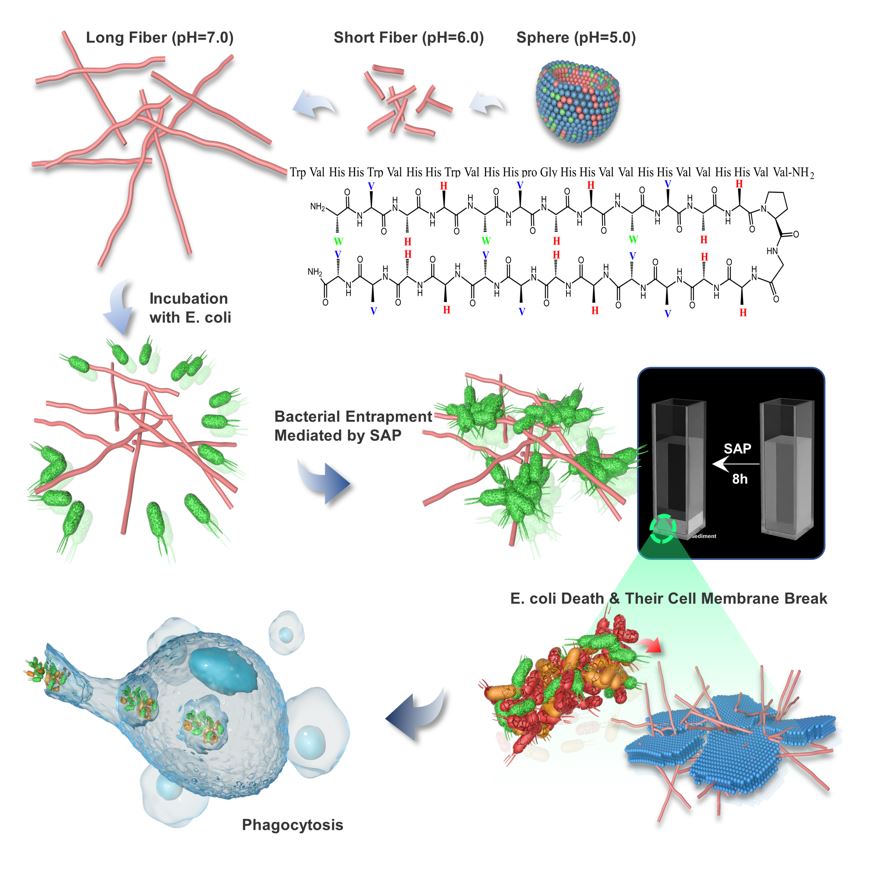
2.1 Design Principles and Molecular Basis of SAP
The SAP was established by considering specific amino acids with special properties and the diagonal cation-π interaction. SAP contains aliphatic or aromatic amino acids (9 Val and 3 Trp) and basic amino acids (12 His) as a pH switch providing multiple interaction capabilities at different pH values in the supramolecular self-assembly system. It was reported that His contains an aromatic imidazole side chain, which provides cation-π (when His is protonated, His+), π-π stacking, hydrogen-π, and hydrogen bonding with other molecules or coordination interactions with metallic cations to play a significant role in protein-protein interfaces and catalysis[25] . Under the acidic condition, the side chains were protonated, which increased the positive charges of the SAP molecules, prompting them to attract with the anionic bacterial membrane. Meanwhile, His, as a pH switch, changed the hydrogen bond energy of self-assembly system when it was protonated and triggered the microstructural transformation of self-assembled peptide. Under the neutral condition, the imidazole ring deprotonated and could be used as a hydrogen donor and acceptor, assisting SAP molecule aggregation into supramolecular structures. In this study, acidic (pH=5.0 and pH=6.0) and neutral (pH=7.0) conditions were selected as model environments for investigating the transformation of nanostructures and biological functions. Besides, the amino acid Trp, containing an aromatic ring, which is reported to serve as hydrogen bond donors and acceptors, facilitates the formation of hydrogen bonds[19], provides hydrophobicity strength for assisting with the formation and stability of nanostructures via π – π stacking and plays an important role in acceleration and stabilization of amyloidogenic assemblies[10]. Meanwhile, Trp-rich peptides act on the bacterial cytoplasmic membrane[26], interact strongly with hydrophobic phospholipid tails, and facilitate peptide insertion into the phospholipid bilayer for permeabilization or pore formation[27]. The hydrophobic amino acid Val was selected because of its strong propensity for forming intermolecular hydrogen bonds, as Val typically possessing high β-sheet forming propensities, which easily leads to the formation of fibers[28]. Furthermore, Trp and His were adapted in two consecutive non-H-bonded sites participating in a diagonal cation–π interaction, which provided driving force to maintain and stabilize the β-hairpin structure[29,30]. Then, equal amounts of His and Val were distributed to other sites to possess enough positive and hydrophobic properties for antimicrobial activity. Additionally, one of the type II’ β-turns, DPro–Gly, was selected as a linker to connect two strands to form a β-sheet structure. The C-terminus of the peptides was aminated to further enhance stabilization (Fig. 1 a).
2.2 pH-triggered Microstructure Transformation of SAP
The self-assembling ability and the critical aggregation concentration (CAC) were investigated via a 1-anilino-8-naphthalene sulfonate (ANS) fluorescence assay, which is an anionic dye that is not fluorescent in water but has high fluorescence intensity while binding to macromolecules. The CAC was determined as described by Fung et al, the ANS fluorescence intensity values at 475 nm were selected, the profile with different peptide concentrations was fitted with two straight lines, and the peptide concentration of intersection indicated the CAC[19]. The results were shown in Fig. 1 b. The ANS fluorescence intensity increased with increasing peptide concentration and aggregation strength at pH 5.0, 6.0 and 7.0, indicating that the SAP molecule could aggregate to form a supramolecular organization under all three pH environments. The CACs of the SAP were shown in Fig. 1 c. SAP was able to assemble above concentrations of 1.0 μg/mL,10.5 μg/mL and 13.24 μg/mL at pH 5.0, 6.0 and 7.0, respectively, demonstrating that the lower the pH was, the greater the aggregation ability of SAP molecules.
The microstructure of SAPs in different environments was investigated via transmission electron microscopy (TEM). The TEM results showed that SAP at a concentration of 15.6 μg/mL self-assembled into spherical micelles at pH 5.0 (Fig. 2d). Spherical micelles transformed into nanofibers after adjusting the pH to 6.0 and 7.0. Compared with neutral conditions, the nanofibers were shorter at pH 6.0, which indicated that the microstructure of the SAP gradually transformed from spherical to nanofibers with the change in pH conditions, and as the pH increased, the microstructure of the SAP transformed into longer nanofibers. Then, the secondary structure transition of the SAP was investigated using circular dichroism (CD) spectroscopy under different pH conditions. As Fig. 1e shown, the spectrum of the SAP at pH 5.0 had a negative band at 200 nm indicating that the peptide adopted a random coil configuration[31], while increasing the pH, it showed a positive peak at 199 nm and a strong negative peak at 218 nm, indicating that the peptide adopted a large amount of β-sheet structure due to the increase in intramolecular and intermolecular hydrogen bonding among the peptide[32]. These data were in direct agreement with the transition of the microstructure when changing the pH environment. Below the acidic condition (pH=5.0), the side chains of His were protonated and positively charged aromatic His residues changed hydrogen bonding energies between the residues and prevented the formation of rigid β-sheets, thus the SAP adopted a random coil configuration. The stronger hydrophobic interactions provided by Trp and Val promoted the SAP to form micelles of finite size, which followed a closed association pathway above a critical micellization concentration. With the increase in the pH (from acidic to neutral pH), the imidazole ring was protonated, which could be either in the neutral or positively charged form, involving in the conformation of hydrogen bonding interactions as hydrogen bond donors and acceptors[33]. Increasing the hydrogen bonding energy promoted the SAP to follow an open association scenario, resulting from the step-by-step aggregation of molecules to form board distribution of β-sheets. The bilayer was formatted with the hydrophobic faces of two layers, which were buries onto each other to separate the hydrophobic moieties from water[34]. And with the formation of hydrogen bonding interactions between adjoining hairpins, fibril structures of the SAP were formed and extended the length of fibers with the increase of pH[35].
A thioflavin T (ThT) fluorescence assay also confirmed this conformational transformation of SAP. The fluorescence values of the peptide were the lowest at pH 5.0 and increased with increasing pH, indicating that the peptide gradually formed an amyloid protein fiber structure with increasing pH (Fig. S3a). The hydrodynamic sizes of the SAP micelles at pH 5.0 were evaluated by dynamic light scattering (DLS) using a Zetasizer Nano ZS90, which presented hydrodynamic radii of ~ 119 nm (Fig. S3b). The measured ζ-potential of the SAP nanoparticles decreased from +26.2 to +1.83 mV upon an increase in the pH from 5.0 to 7.0, which indicated that SAP fibers tended to transform to a neutral status (Fig. S3c).
2.3 Antimicrobial Activity, Antimicrobial Mechanism and in vitro toxicity of SAP
The antimicrobial activity of the self-assembled peptide was tested by colony counting experiments against clinically prevalent bacterial species, including Escherichia coli, Salmonella typhimurium, Listeria monocytogenes, and Bacillus cereus. After treatment of bacterial cells ((2−5) × 105 CFU/mL) with the peptide at different concentrations ranging from 3.9 μg/mL to 250 μg/mL for 3 hours, 50 μL of the bacterial solution was further continuously diluted, and a 10 μL aliquot from each dilution was plated on TSB agar plates. As shown in Fig. 2a, the SAP displayed remarkable antimicrobial activities under acidic conditions above the CAC. Especially for the representative strain E. coli ATCC 25922, the SAP at the highest concentration tested (250 μg/mL) exhibited highly effective activity, which reduced survival by approximately three orders of magnitude at pH 5.0 and two orders of magnitude at pH 6.0. The same results were observed in antibacterial experiments against S. typhimurium ATCC 14028, in which 250 μg/mL SAP reduced the survival by approximately two orders of magnitude and one order of magnitude at pH 5.0 and 6.0, respectively. For gram-positive bacteria Listeria monocytogenes CGMCC 1.10753, 250 μg/mL SAP reduced two orders of magnitude at pH 5.0 and 6.0. For Bacillus cereus CGMCC 1.932, 250 μg/mL SAP reduced one and two orders of magnitude at pH 5.0 and 6.0, respectively. On the contrary, no detectable antibacterial activity against bacteria was exhibited at various concentrations at pH 7.4 (Fig. 2a). The above results showed that the formation of nanostructure is indispensable for SAP to inhibit the bacterial proliferation, and the lower the pH was, the better the antibacterial effect of SAP was, indicating that spherical micelles could effectively inhibit the bacterial growth and that the antibacterial effect decreased gradually with the transformation from spherical nanoparticles to fibers. The shape of the nanostructure influences the antimicrobial properties, and spherical nanoparticles are more effective than fiber nanoparticles[28]. Additionally, the net charge of the nanoparticle might contribute to the altered antibacterial activity. The electrostatic interaction between cationic AMPs and anionic bacterial membranes is considered important to the driving force for bacterial-peptide adsorption[41,42]. Under acidic conditions, His was protonated, which enhanced the positive electricity of the nanoparticle, increasing the electrostatic adsorption of peptides and the bacterial surface[38]. Moreover, we discovered an interesting phenomenon in the course of the antibacterial experiment that bacterial clusters could be observed after treatment with SAP, which indicated that SAP could entrap bacteria and promote bacterial agglutination, and we would explore additional experiments to investigate this interesting finding, subsequently.
In addition, self-assembled molecules have been reported to overcome proteolytic degradation challenges. To evaluate the properties of SAP, peptides were incubated with trypsin and chymotrypsin at mass ratios of 2:1, 1:1, 0.5:1 and 0.25:1 (enzyme: peptide for 1 hour at 37°C and then the antimicrobial activity of the hydrolyzed peptide was assessed. The results were shown in Fig. 2b, SAP maintained its antibacterial activity after incubation with trypsin compared with unhydrolyzed peptide, even when the mass ratio of enzyme to peptide was 2:1 (p > 0.05), suggesting that SAP could resist the hydrolysis of trypsin. This is because trypsin preferentially cleaves at basic residues (Arg and Lys) rather than His[38]. And there were no significant differences, despite the slight increase in the number of colonies of E. coli after treatment with the hydrolyzed peptide compared with after treatment with the unhydrolyzed peptide (p > 0.05), indicating that nanoparticles were slightly hydrolyzed by chymotrypsin in a concentration-dependent manner. This may be due to Trp contained in the SAP, which is prone to digestion by chymotrypsin[36]. However, melittin, which was used as a control, was completely hydrolyzed by trypsin and chymotrypsin after incubation for 1 hour and became inactive against E. coli even at the maximum tested concentration (250 μg/mL). This is because melittin contains Leu, Trp, Lys and Arg, which are prone to hydrolysis by chymotrypsin and trypsin (Fig. 2b).
To assess the salt sensibility of the SAP in vitro, the antibacterial activity in the presence of NaCl was monitored using a viable colony count assay against E. coli ATCC 25922 as the model strain. As shown in Fig. 2c, there was a slight increase in the number of colonies of E. coli after treatment in the presence of NaCl, but no significant differences were detected in the treatment of SAP above 15.6 μg/mL in the presence of 100 mM NaCl and above 125 μg/mL in the presence of 150 mM NaCl, compared with the control in the absence of NaCl, suggesting that the presence of NaCl could impact the biological function of the nanoparticles. This may be due to the effects of the electrostatic interaction between antimicrobial peptides and bacterial membranes in the presence of Na+[39]. Moreover, NaCl also showed a protective effect on cell survival at pH 5.0 and there was no significant difference after treatment in the presence or absence of NaCl above 31.2 μg/mL (Fig. S4)[40].
AMPs are considered less likely to develop drug resistance because they could compromise the integrity of microbial cell membranes, thereby evading the pathways by which bacteria develop rapid antibiotic resistance[41]. Based on the above results, pH 6.0 was selected as the model condition to investigate the antibacterial mechanism because under this pH, the SAP showed antibacterial activity and the E. coli showed substantial growth improvement. Membrane depolarization and permeation were evaluated using a 3,3′-dipropylthiadicarbocyanine iodide (diSC3-5) fluorescence assay. DiSC3-5, a membrane potential-sensitive dye, displays increased fluorescence upon changing the cytoplasmic membrane potential. Fig. 2d showed that rapid dose- and time-dependent increases in the fluorescence intensity induced by various concentrations of SAP, as well as melittin, a typical membrane disruption peptide. Taken together, these results demonstrated that SAP could lead to membrane potential perturbation and pore formation. Scanning electron microscopy (SEM) and TEM were used to evaluate the effect of SAP treatment on bacterial morphology and intracellular ultrastructural alterations at the highest test concentration, respectively. As shown in Fig. 2e, in comparison with the control, the E. coli membrane induced by treated with 250 μg/mL SAP at pH 6.0 was seriously damaged, while SAP-untreated E. coli showed a complete membrane surface. Numerous nicks and tears were evident in the cell membrane of the 250 μg/mL SAP-treated E. coli, where membrane disintegration and clumping were distinctly observed and debris and lysis of the cells were also evident. For intracellular ultrastructural alterations of E. coli after treatment with SAP, Fig. 2e showed the obvious separation of the cytoplasmic membrane and outer membrane and disruption of the outer membrane, which caused the formation of obvious cavities within the E. coli, determined that bacteria lost the intracellular content via the pores after treatment with nanopeptides in an acidic environment. In contrast, enriched cellular content and complete outer and inner membranes were observed in E. coli without nanopeptide treatment. According to the SEM and TEM and diSC3-5 results, the distinct morphological disruption caused by SAP illustrated that the bacterial membrane was an important target of the antibacterial activity of nanopeptides in an acidic environment.
In addition, biocompatibility assessments of peptide are the prerequisite for further application. Thus, for testing the application potential of SAP, RAW264.7 cells, HEK293T cells and mouse red blood cells were selected as model cells for evaluating the safety of SAP at pH 5.0. 6.0 and 7.0. The peptide cytotoxicity against RAW264.7 and HEK293T cells was tested via a 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT) dye reduction assay. For RAW264.7 macrophage cells, the results showed that the cell survival rate was maintained above 80% after treatment with the peptide at pH 6.0 and 7.0, even at the highest test concentration of 250 μg/mL, but at pH 5.0, the cell survival rate showed a concentration-dependent reduction, which was approximately 70% after treatment with 250 μg/mL SAP for 4 hours (Fig. 3a). This result was sharply in contrast to the results obtained for melittin, in which the cell survival rate decreased to 30%, 25% and 10% after treatment with 7.8 μg/mL melittin at pH 5.0, 6.0 and 7.0, respectively (Fig. 3b). Similar to RAW264.7 cells, the survival rate of HEK293T cells induced by the SAP was above 80%, and no significant differences were observed in the cell survival rate under the three pH conditions (Fig. 3c). The cell survival rate induced by 7.8 μg/mL melittin decreased to 60%, 40% and 25% at pH 5.0, 6.0 and pH 7.0, respectively (Fig. 3d).
In the hemolysis assay, mouse erythrocytes were added into different concentrations of the peptide in three environments and the toxicity was determined by the amount of hemoglobin released by the lysis of erythrocytes. The results were shown in Fig. 3e, the SAP induced no hemolysis against mouse red blood cells at pH 5.0, 6.0 and pH 7.0, not exceeding 5% even at highest concentration 250 μg/ml, but 62.5 μg/ml melittin caused hemolysis exceeding 50% at pH 5.0, 6.0 and 7.0, respectively (Fig. 3f).
Based on the results of cytotoxicity and hemolysis assay, spherical nanoparticles showed higher toxicity on cells than nanofibers, indicating that the transition of peptides to fibers reduces the interaction with the membrane, which was validated by the antimicrobial activity results. Moreover, compared to the natural peptide melittin, the SAP showed high biocompatibility and fewer side effects, which indicated that the SAP has the potential for clinical use as a promising therapeutic.
2.4 Bacterial Agglutination and Phagocytosis Mediated by SAP
Based on phenomenon that SAP could promote bacterial agglutination in the antibacterial assay, bacterial agglutination assay was performed to assess the entrapment function (Fig. 4a), which was evaluated by the turbidity of the bacterial solution and the number of colonies forming units of bacterial supernatant after SAP treatment as Phoom et al. described[24]. To investigate the aggregation ability of SAP in different environments, 250 μg/mL SAP, the highest concentration tested was introduced into E. coli ATCC 25922 culture (108 CFU/mL) prepared in an EtOH-sterilized cuvette. As shown in Fig. S5, bacterial clumping was observed under three pH conditions after adding SAP, and these clumps sedimented to the bottom of the cuvette within 8 hours. In particular, the supernatant of the bacterial solution was substantially clear after 3 hours at pH 6.0, and the SAP somewhat reduced the turbidity of the bacterial supernatant at pH 5.0 but not as fast that at pH 6.0 (Fig. S5a and b). In a neutral environment (pH=7.0), the SAP could entrap bacteria to form milky bacterial solutions, indicating that peptides and bacteria quickly form a higher degree of aggregation (Fig. S5c). From the results of the change in bacterial numbers in supernatant, the sedimentation rate of bacteria was the fastest in pH neutral solution, and the number of colonies was significantly decreased from 108 CFU/mL to zero within 3 hours, followed by pH=6.0 from 108 CFU/mL to zero within 8 hours (Fig. 4b). In contrast, agglutination and sedimentation were not observed for the untreated control. These results revealed that the pH-triggered self-assembled peptide SAP has the ability to recognize and trap bacteria, and the higher the pH was, the faster the bacteria clumped. To determine whether the bacterial sedimentation was due to the automatic sedimentation of the peptide by changing pH, 250 μg/mL SAP was added into 4-(2-hydroxyethyl) piperazine-1-ethanesulfonic acid (HEPES) buffer (pH=5.0, 6.0 and 7.0) for 8 hours. Indeed, incubating the SAP without E. coli did not aggregate as previously identified after the addition of E. coli, indicating that amyloid-like aggregation is triggered upon incubation with bacterial cultures (Fig. S6).
Considering the antimicrobial and agglutination properties, pH=6.0 was selected as the model environment to investigate the bacterial agglutination efficiency of different SAP concentrations (3.9-250 μg/mL). As shown in Fig. S7, at SAP concentrations above 62.5 μg/mL, obvious formation of aggregation could be observed. In particular, the formation of aggregation was observed within 0.5 hours at peptide concentrations of 250 μg/mL and 125 μg/mL, and at peptide concentrations of 62.5 μg/mL, the formation of aggregation was observed within 1 hour. However, at concentrations below 31.2 μg/mL, no significant bacterial aggregation was observed, even though the peptide showed antibacterial activity above 15.6 μg/mL. This was also illustrated by the results of the change in bacterial numbers in the supernatant, and a significant decrease was observed in the number of colonies when the peptide concentration was above 62.5 μg/mL. However, no aggregation phenomenon was observed in the concentration of peptide in the range of 15.6 μg/mL to 31.2 μg/mL. Subsequently, bacterial aggregation mediated by SAP was observed using fluorescence microscope. After treatment with 250 μg/mL SAP, E. coli particles aggregation and internalization of nucleic acid stain PI into E. coli was observed, that determined SAP could induce bacterial aggregation and accompanying bacterial death (Fig. 4d).
This bacteria-entrapped function of peptide was first reported by Bevins and co-workers in 2012, they seminally investigated that the human α‑defensin 6 (HD6) undergoes ordered self-assembly into fibrils and nanonets to entrap and agglutinate Salmonella Typhimurium to prevent human gastrointestinal pathogen from invading intestinal mucosal cells, rather than kill bacteria directly[22]. This mechanism is also observed in neutrophil extracellular traps (NETs), an extracellular web-like neutrophil-derived protein network, which plays a role to trapping invading bacteria such as Klebsiella pneumoniae and inducing a series of immunoreaction[42]. Focused on the host defense model of HD6, Phoom and co-workers investigated the molecular basis of self-assembly behavior and capacity of entrapping bacteria via site-directed mutagenesis[24]. Absent of bacteria and other biomolecules, four HD6 monomers could form a hydrophobic pocket and the reciprocity of these tetramers promote the formation of fiber chains in aqueous buffer[43]. Among these reciprocities, the hydrophobic interaction provided by Phe2 and Phe29 is essential for the self-assembly ability and the Phe2Ala and Phe29Ala variants caused the depolymerization of HD6 and failed to prevent L. monocytogenes invasion, which indicates that the self-assembly is important for microbial entrapment[24]. The experimental results of this study further verified this point. The SAP could trap E. coli at a concentration of 62.5 μg/mL, this was above the CAC under three environments. In the neutral, the higher turbidity was observed in the bacterial solution due to the higher aggregation degree and instant formation of larger bacterial clusters, and that was because SAP was in a neutral status, and the hydrophobic force of Val and Trp mediates the interaction between peptide and bacteria, inducing a rearrangement of peptides, promoting the agglutination of bacteria via aggregation of surface-attached peptide molecules[21,50]. Moreover, different bacteria seem to be developed different nanonet structures and the amount of nanonets wrapping via microscopic analysis[45]. And Chu and co-workers investigated that the interactions exist between the HD6 nanonets and the surface protein of S. Typhimurium. These evidences indicate that this entrapment seem to be mediated by the interactions between peptide and bacterial surface protein or other components, and Yu and co-workers determined that this possibility is feasible. They introduced a ligand peptide, which can specially bind to lipoteichoic acid (LTA), into the self-assembly system and the specific binding to LTA mediated the transformation of nanoparticles into nanorods, accompanied by the agglutination of S. aureus[23]. As for our study, further studies are needed to determine whether SAP-mediated bacterial agglutination is related to peptide-receptor interactions. In addition, Lim and co-workers also attributed this bacterial-peptide aggregation phenomenon to ligand-receptor interactions and pointed out that the length of nanofibers has a significant influence on the formation efficiency of bacterial clusters. The longer fibers tended to promote agglutination of large bacterial clusters, which seem to have more bacterial binding sites, but the short nanofibers seem to interconnect a single or small number of bacteria[46]. And our results also confirmed this point that longer fibers could cause higher agglutination degree of E. coli. It is noteworthy that in our study, the SAP had antimicrobial activity in addition to its trapping function. Marc and co-workers reported that the aggregation process triggered the subsequent membrane permeabilization[21], but our results were somewhat at odds with this point. The SAP-mediated bacterial agglutination degree observed in acidic conditions was weaker than in neutral conditions, but bacterial aggregation process should not exclude the possibility that parts of bacteria were killed directly by the peptide. The antimicrobial activity of peptide is determined by a positive charge and hydrophobicity of peptide, while the trapping process is mediated by hydrophobic interactions between peptide and bacterial surface. The possibility of these two processes occurring simultaneously cannot be excluded, but they are independent of each other.
Subsequently, we hypothesized that bacterial agglutination could promote the effective clearance of bacteria particles of phagocytes at bacteria-infected sites. To test this, the uptake of E. coli particles by RAW 264.7 was measured using 3D-SIM super-resolution microscopy. As shown in Fig. 5e, E. coli particles, aggregated by SAP, increased the internalization of RAW 264.7, compared with control, which showed limited phagocytic activity of RAW 264.7 cells. The result indicated that SAP are able to aggregate bacteria and promote phagocytic uptake, which play an important role in clearance of E. coli by phagocytes.
2.5 In-vivo efficacy and biocompatibility of SAP
The above results of bacterial agglutination assay indicated that SAP showed the best entrapment activity in neutral environment. For further evaluate in vivo therapeutic efficacy of SAP, a peritonitic-bacteremic mice model was implemented to explore the entrapment ability in vivo to prevent bacteria from invading the animal tissues. As shown in Fig. 5a, each mouse in the infection group was intraperitoneally injected with 1.5× 108 CFU/mL E. coli ATCC 25922 and each mouse in the control group was injected with the same amount of saline. At 1 h postinfection, each mouse in the treatment group was intraperitoneally injected with 10 mg/kg SAP and 1 mg/kg colistin. Compared with control, bacterial burdens in liver, spleen, kidney, and peritoneal cavity were significantly reduced after the treatment of SAP and colistin (p<0.05), and there are no significant differences only in organs between SAP and colistin treatment (p>0.05) (Fig. 5b), which indicated that acidic peptides initially exhibit antimicrobial activity like colistin, resulting in a decrease of the number of bacterial colonies in the blood and the peritoneal cavity, and then the peptides translated to a neutral state that entrapped bacteria and prevented them from invading the tissue. Moreover, the levels of proinflammatory cytokine TNF-α, IL-6, and IL-1β of SAP treatment were significantly lower compared with control (p<0.05), and the levels of anti-inflammatory cytokine IL-6 and IL-10 of SAP treatment were significantly higher compared with control (p<0.05) and no differences were noted compared colistin treatment (p>0.05) (Fig. 5c). The H&E staining showed that the treatment of SAP and colistin alleviated E. coli-induced tissue damage, such as hepatocytes damage and loss of the nucleus and cytoplasm, swelling of the glomerulus, and lymphocyte infiltration in the spleen, which promoted liver cell regeneration, normalization of glomerular structures, and spleen tissues (Fig. 5d). These data indicated that SAP could attenuate the bacteremia-induced inflammatory cytokine levels and organ tissue damage via killing and entrapping bacteria and regulating the body’s immune system. In a word, in-vivo and in-vitro biological activity, biocompatibility and efficacy highlight the promoting potential of pH-triggered β-hairpin forming nanopeptide for the treatment of acid-tolerant enteric pathogens induced infection.
For further investigation of the safety of SAP in vivo, 6 mice in each group were injected intraperitoneally with 10 mg/kg, 20 mg/kg, 40 mg/kg SAP, and body weight and relative organ weight were monitored to assess the toxicity of SAP, and then serum was taken out to measure serum alanine aminotransferase (ALT), alkaline phosphatase (ALP), total blood bilirubin (TBIL) activity, blood urea nitrogen (BUN) and serum creatinine (CRAE) for assessment of the hepatotoxicity and nephrotoxicity, respectively[47]. As shown in Fig. 6, there were no significant differences in the body weight (Fig. 6b), and the liver, kidney and spleen index (Fig. 6c) of all groups. Meanwhile, no significant differences were noted on the levels of ALT, ALP, TBIL, BUN and CRAE between the control group and peptide-injected groups (Fig. 6d and e). Additionally, histological examination (H&E) also showed that there are no obvious tissue abnormalities of all treatment groups (Fig. 6f). Taken together, SAP have good biocompatibility, resulting in a wide range of development and application potentials.
{kind=link}
{kind=link}