Filariasis, and anthroponotic onchocerciasis have been reported worldwide and have caused problems either in animals or humans [5, 19], as many cases of zoonotic onchocercosis are reported [20]. Anthroponotic onchocerciasis has also been documented in Africa and Central/South America and has resulted in visual impediments in the affected individuals [6, 21].
In the current study, parasitic nodules in the form of yellowish subcutaneous nodules in various parts of the body have been observed in Japanese Sika deer (Cervus nippon) captured from Gifu and Shiga Prefectures in Japan. These findings are in agreement with the previously reported onchocerciasis in red deer (Cervus elaphus) in Switzerland [20]. The morphology of the present filarioids have the main characteristics of the genus Onchocerca that given by Bain [22] i.e. females have a thickened cuticle with transverse annular ridges on their surface and weakly developed muscular layer.
Histopathological and the measurements analyzed, matched with those given in the description of O. flexuosa, carried out by earlier investigators [7, 8, 23]. The detected long female of O. flexuosa is identical to the female of O. jakutensis discovered by Bosch et al [20] from the Swiss red deer. Chronic parasitism in the form of multiple granulations around the worm and marked leukocytic cell infiltration of eosinophils, lymphocytes, macrophages, and plasma cells were observed and these granulomata were found mainly around female worms. Presences of many degenerated worms within several cavities in the nodules are an indication of a strong immunological response against O. flexuosa [8, 24]. Female worms were found more frequently in the nodules than males, they were embedded in the nodular fibrous tissue, and most of them were usually immobile due to an early degenerative alteration of their muscles. In contrast, mobility activity of males is higher than females as they can migrate subcutaneously leaving the nodules. Similar conclusions have been reached before [8, 25, 26].
The presence of rivers and mountain streams enhance the vectors survival; resulting in high prevalence and intensity [8, 25, 26]. Both temperature and moisture are the most important environmental changes for all organisms. So, the ecology and the epidemiology of O. flexuosa are differing according the location. The prevalence of O. flexuosa in Sika deer was unknown in the Japan especially in the above mentioned examined regions. Prevalence of Onchocerca spp in wild and domesticated animals had previously appeared to be restricted to central and eastern Japan in Myazaki city (unpublished manuscript, gene bank reference number at https://www.ncbi.nlm.nih.gov/nuccore/AP017692.1), perhaps because these were the only areas where they had been studied. To the best of our knowledge, this is the first record of O. flexuosa in that area, as there is only one record of O. flexuosa from Myazaki/Japan in the GenBank.
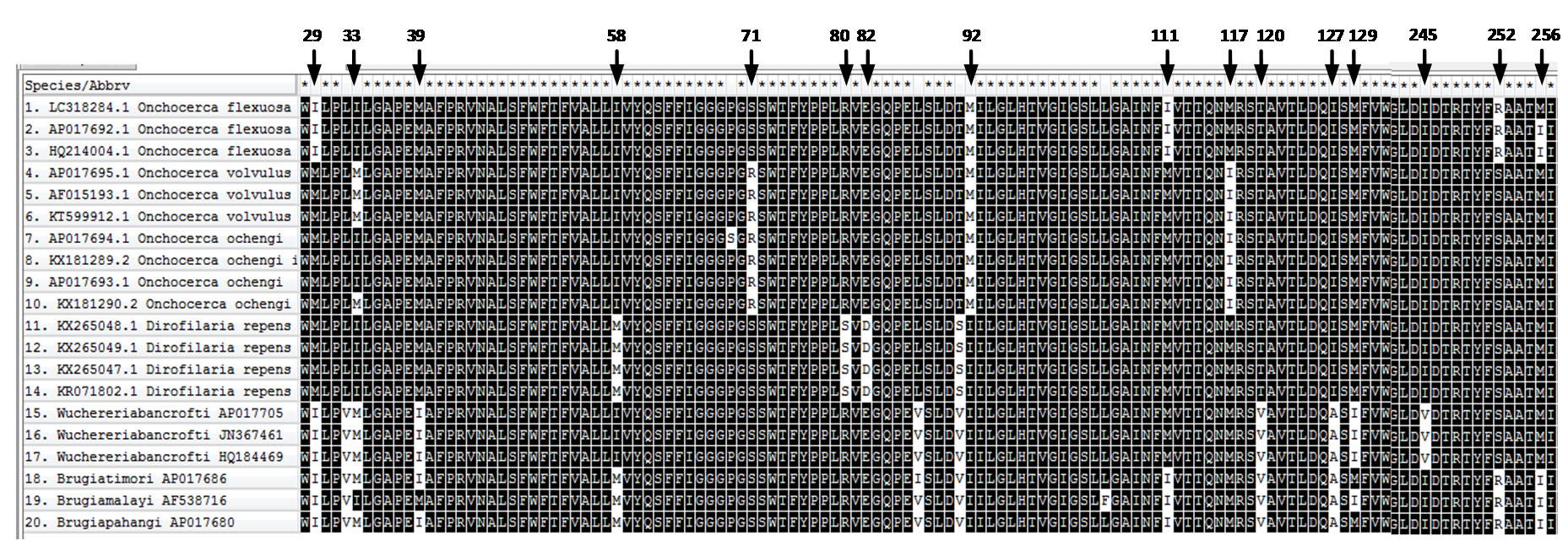
In the present study, in order to confirm the molecular identity of O. flexuosa and to investigate its phylogenetic position, we compared the nucleotide sequences of cox1, NAD2 and ITS2–28S rRNA with those of all Onchocerca species available in GenBank. The nucleotide sequence of cox1 in our samples of O. flexuosa was identical to other O. flexuosa isolated from Japan and was similar (0.2% nucleotide differences) to O. skrjabini O. boehmi and O. lupi. The present isolate of O. flexuosa differed from O. dewittei japonica and O. takaokai n. sp with differences of 4.7–4.5% respectively in the nucleotide sequences of cox1. Sequence analyses of the ITS2–28S rRNA gene of O. flexuosa revealed a 5.8 − 7.4% difference compared with the nucleotide sequences of O. dewittei japonica and a 4.8– 8.5% difference compared with that of other congeners. Similarly, the ND2 sequence analysis revealed a 5.8 − 7.4% nucleotide sequences difference compared with that of O. dewittei japonica and a 4.8– 8.5% difference compared with that of other congeners.
P-distances in nucleotide sequences of examined genes as mentioned in the results and our phylogenetic analysis based upon cox1, NAD2 and ITS2–28S rRNA genes indicated a close affinity between the present isolate of O. flexuosa and the previously isolated O. flexuosa, O. skrjabini, and O. eberhardi, isolated from Sika deer or serows in Japan. In contrast, O. flexuosa was distant from O. takaokai n. sp and O. dewittei japonica. Moreover, our results confirmed the previous reports [27, 28], indicated that the phylogeny of the family Onchocercidae based on the previously mentioned genes sequences plays a vital role in the classification of filarial worms along with the classification based upon morphological characters.
In a previous studies, DNA analysis and barcoding based on cox1 and other mitochondrial genes was considered a reliable method for species identification of filarioid nematodes, with a high coherence with classical taxonomy [29]. In recent studies different approaches to species recognition generated similar results [30], which suggested that integration of traditional and molecular approaches to species recognition is possible [31]. More recently Identification of Onchocerca spp using cox1 as a molecular marker is considered accurate as previously indicated for Onchocerca spp [32], as well as other filarial species [29]. DNA analysis was considered a good and reliable approach for taxonomical identification of filarioid nematodes, with a high coherence with classical taxonomy [29].
In the present study, the filarial parasite was identified as O. flexuosa based on traditional and molecular analysis. Phylogenetic analysis based on cox1, ITS2–28S rRNA, and NAD2 gene sequences revealed that only three main groupings were recorded in the family Onchocercidae, viz., Onchocerca spp, Dirofilaria spp, and Brugia + Wuchereria. Similar conclusion has been reached by [16, 33] based upon molecular characteristics and phylogeny of the Filarioidea superfamily. Thus, the phylogenetic aspects of filarial worms may be resolved by analysis based on cox1, ITS2–28S rRNA, and NAD2 genes.
In our opinion, the light of the present work suggests that cox1, ITS2–28S rRNA, and NAD2 appeared an appropriate molecular marker for identification of filarioid nematodes up to species level. Similar suggestion has been reached by using cox1 and 12S rDNA genes for identification of filarial nematodes up to species level [29]. The phylogenetic analyses of cox1, the ITS2–28S rRNA, and NAD2 genes in that study indicated that O. flexuosa isolated from Japanese Sika deer was identical to Onchocerca spp in terms of nucleotide sequences. According to [29], no overlap of intra- and interspecific nucleotide divergence at the cox1 gene occurs at distance values greater than threshold values (approximately 4.8%). Based upon intra- and interspecific nucleotide divergences, in this study we calculated the genetic distance between cox1 from two different species, and it was greater than the threshold value of 4.8%. Therefore, our findings clearly document the conclusion that the present isolate is O. flexuosa, which is distinguished from O. dewittei japonica, O. takaokai and other congeneric species at the species level, and has a close affinity to other O. flexuosa previously reported.
The sequencing analysis is of great importance to identify the origin of the genus Onchocerca and its relation with other members of the family and this characteristic could be used to medicinally control the infection by targeting a specific gene. The emergence of more infectious diseases is a significant problem for the world economy and public health. Majority of these diseases originate in wildlife and spread to other species including animals and humans, and their incidence has increased significantly over the past few decades [34]. Thus, disease surveillance efforts and accurate diagnosis should be applied frequently especially in wildlife to control spread of these pathogens to human or other species.
{kind=link}
{kind=link}
{kind=link}