In the present study, we isolated three pure strains of Bacillus sp., from the grass carp pond water. Among them, a particular isolate, namely, Bacillus velezensis LG37, was successfully screened using minimal media with ammonia as the sole nitrogen source for the best ammonia nitrogen assimilation efficiency; thus, it was chosen for further analysis.
Growth characteristics of LG37 culture conditions
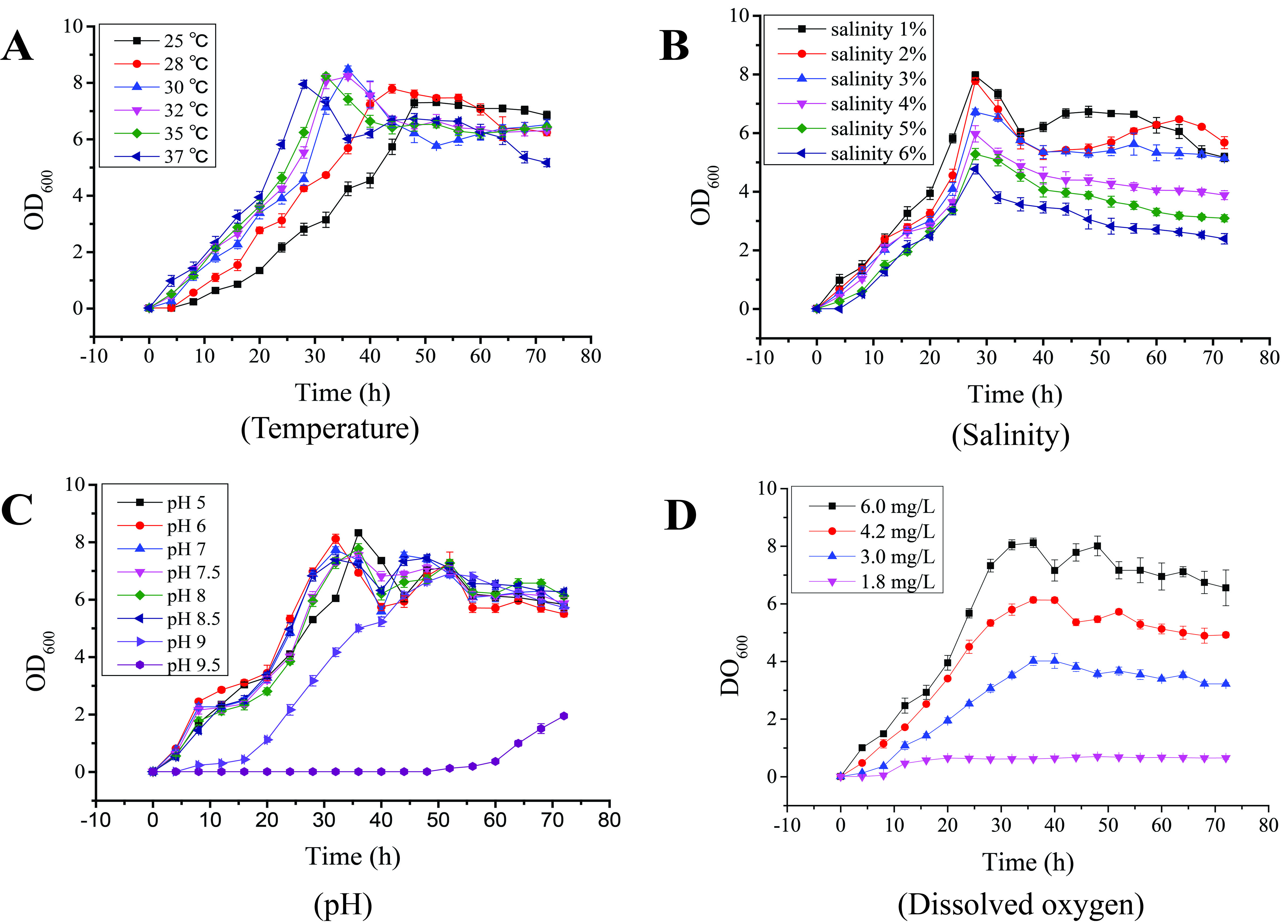
The bacterial growth kinetics of wild-type LG37 was analyzed by culturing in LB broth, and the growth was measured by spectrophotometry. From 25 to 37 °C, the LG37 showed an excellent growth trend. The peak levels OD (≈7) were maintained for almost 30 h in the static cultivation environment (Additional File 1). As shown in Additional File 1, LG37 on 1 - 6 % of salt concentrations had a sharp and stable increase in their growth exceeding OD600 value of 5 at 28 h followed by a gradual decline. Yet, the levels were maintained above 5 up at to 3% salinity, and the decrease in OD was proportional to salt concentration in the extended period. The LG37 growth was normal at pH 6-8.5, while slow growth was recorded at pH 9.5. The dissolved oxygen (DO) concentration available in the medium mainly influences the growth of the bacterial population [43]. The increasing growth values were observed in all the DO reactors from 0 to 40 h, and the LG37 growth was proportional to the level of DO. Peaked OD of 8.2, 6.1, and 4 – were obtained for LG37 with initial DO of 6.0, 4.2, and 3.0 mg/L, respectively, at 35 h of the culture period. Conversely, there was no substantial growth recorded in the 1.8 mg/L DO (Additional File 1).
Optimization of Glutamine and ammonia nitrogen concentrations for LG37
Growth characteristics of LG37 were determined at various levels of NH4+ and Gln by OD600. The results showed, with the increase of Gln concentration, LG37 displayed an inclination of growth respect to the amount of Gln (Fig. 2A). The growth curve of LG37 in minimal media with NH4+ (5, 10, 15, 20, 25, and 30 mmol/L) as the sole nitrogen source showed a similar trend to that of Gln, but no significance was observed at 20 - 30 mmol/L NH4+ concentrations (Fig. 2B). However, the growth of LG37 was more significant in the increasing levels of nitrogen; the other nutrients in the minimal media might be a limiting factor for their growth above 20 mmol/L. Given that and to avoid overloading, we preferred an average volume (10 mmol/L) of N-source for both Gln-N and NH4+-N in our supplementary studies.
Transcriptome assembly and functional annotation
The mRNA of the LG37 cells acquired from the treatment group (NH4+-N) and control group (Gln-N), were sequenced by Illumina HiseqTM 2500 to obtain the overview of the gene expression pattern. After removing low quality (Q > 20) ambiguous reads from the raw data, a total of 19091060, 19270408, and 19079520 clean reads from the LG37-Gln (Gln-N1, Gln-N2, Gln-N3), and 18922170, 18818298, and 18865012 clean reads from LG37-NH4+ (NH4+-N1, NH4+-N2, NH4+-N3), were obtained, respectively. The details of GC contents, valid ratio, raw reads, and clean reads were summarized in Additional File 2. We aligned our LG37 data with the known reference genome (B. velezensis FZB42; Accession No: NC_009725.1) using the BWT algorithm to interpret the clean read sequences. A total of 2569 genes from LG37 were annotated as protein-coding genes; additionally, we also predicted 63 new genes, including 30 sense transcripts and 33 antisense transcripts, 2131 for 5’UTR, 2037 for 3’UTR, and 759 predicted multi-gene operons. Summary of alignment with protein-coding genes, predicted transcripts, and predicted RNAs (antisense) were presented in Additional File 3.
The LG37 DEGs potentially intricated in nitrogen assimilation were determined by conducting the statistical analysis through expression > 1.5 fold change and significant difference of q-value < 0.05 as standards. Expression variances were compared between the treatment and control groups using the standardization of RPKM and UQ values. In total, 812 DEGs (76 upregulated genes and 736 down-regulated genes) were screened. Among them, 56 genes met the differential expression genes (DEGs) criteria (i.e. expression value >= 2-fold and p-value <= 0.05). Subsequently, the genes were compared with NCBI and UniProt database for screening the candidate related genes for N-metabolism, resulted in 27 candidate genes, including 18 upregulated DEGs, and 9 downregulated DEGs, listed in Additional File 4.
The regulatory function, the expression pattern in various cellular compartments, molecular function, and biological processes of these genes in treatment and control nitrogen groups of LG37 were overviewed by mapping unto GO and KEGG databases. The results demonstrated that 812 unique proteins assigned to 3,817 GO terms; 1,698 unigenes mapped to biological processes, 860 unigenes mapped to molecular functions, and 1,259 unigenes mapped to cellular components (Additional File 5). In the biological process subclass, the top 3 categories were “cellular process (464), metabolic process (420), and single-organism process (340)”. In molecular function, the top 2 subclass were “catalytic activity (383) and binding (351)”, accordingly, in the cell compartment, the most abundant categories were “cell (411), cell part (411), membrane (163), and membrane part (142)” (Fig. 3). Moreover, the KEGG pathway analysis indicated the DEGs were significantly (p-value 0.001) enriched in five KEGG pathways, comprising pathways associated with Biosynthesis of secondary metabolites (108), Ribosome (36), Biosynthesis of amino acids (51), Carbon metabolism (46), and Photosynthesis (7) (Fig. 4, Additional File 6).
RT-qPCR verification of selected genes
The Illumina sequence of LG37 expression profile data was verified through randomly selected 8 DEGs that including, 5 up-regulated and 3 down-regulated genes (Additional File 7) using RT-qPCR. The results exhibited a similar expression tendency but with slight variation in their levels, which confirmed the reliability of DEGs from the transcriptome sequencing results (Fig. 5).
GlnK is critical for NH4+ assimilation
To understand the functions of the specific genes associated with NH4+ assimilation pathway that short-listed by transcriptome data, especially the significantly upregulated glnK (4.26) and glnL (4.12), were primarily analyzed by RT-qPCR following cultivation of wild-LG37 at different concentrations of NH4+. The glnK and glnL showed a significant increase in their expression with the rise of NH4+ concentration, and the glnK showed a greater tendency than the glnL (Fig. 6A). These results demonstrate that the two-component regulatory systems glnK/glnL play a synergistic regulation function in the LG37 NH4+ metabolism.
Functional analysis of related genes using OEglnK and ΔglnK in NH4+ metabolism
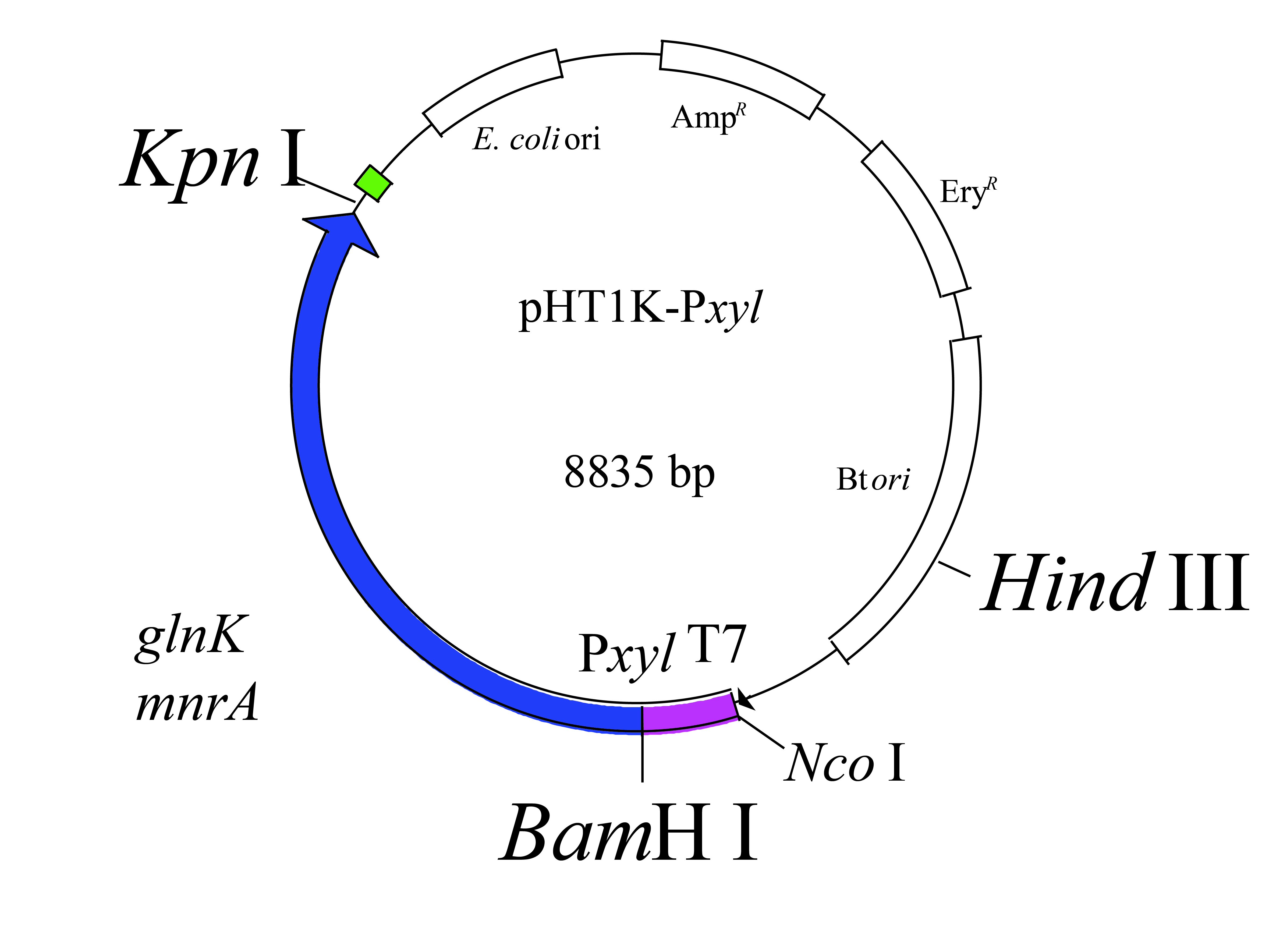
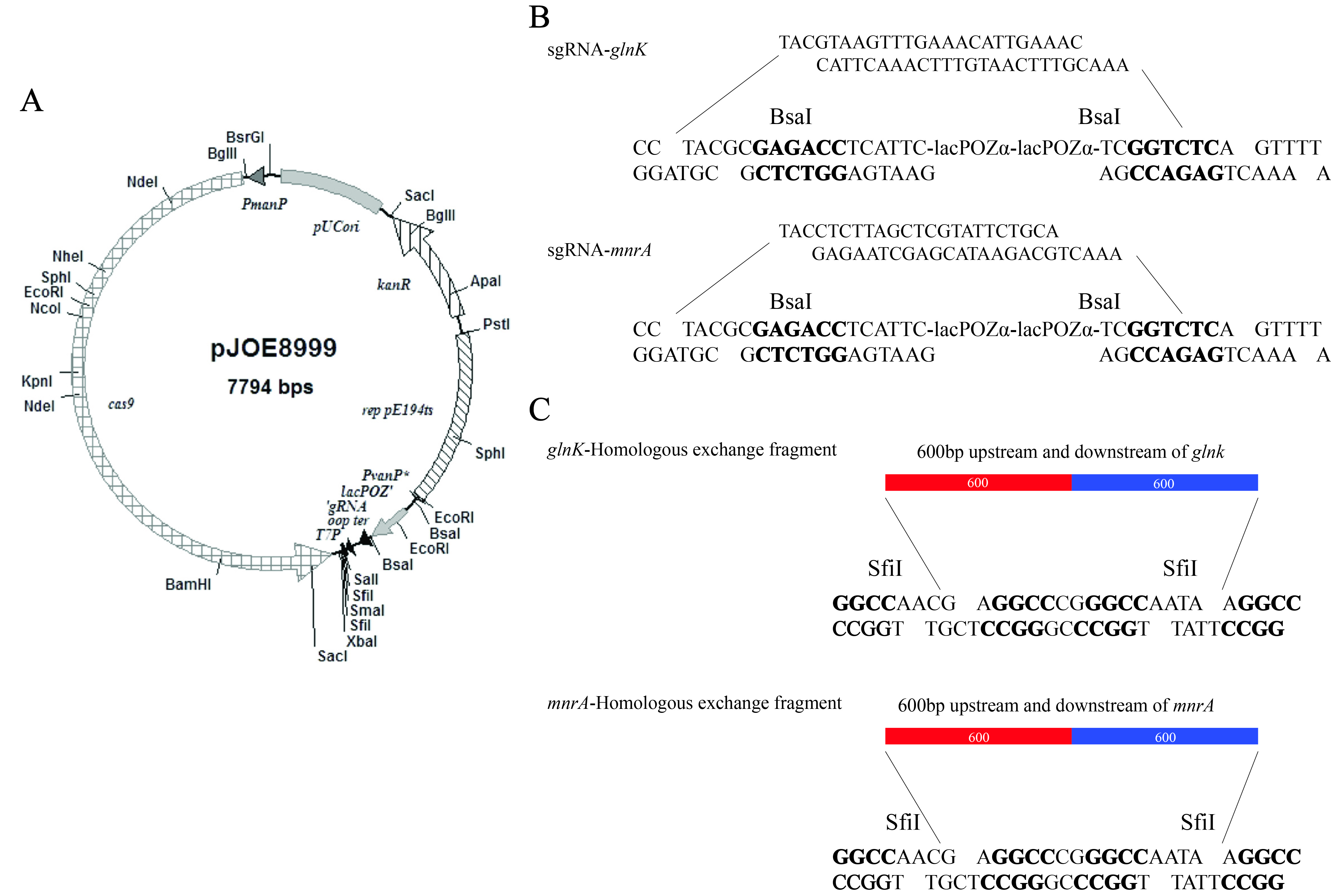
To demonstrate the functionality of the GlnK in NH4+ assimilation pathway, glnK was knocked out (LG37-ΔglnK) by applying the CRISPR/cas9 technology (Additional File 8) using pHT1K plasmid that developed overexpression mutant (LG37-OEglnK) strain (Additional File 9). The mutants were analyzed for the regulation of ammonia assimilation related genes (glnL, amtB, and glnA) by RT-qPCR assays along with wild LG37 strain, following cultivation in 10 mmol/L NH4+ containing minimal medium. The LG37-OEglnK strain increased the expression of about 2.5 folds and 1.8 folds, while the LG37-ΔglnK strain lead to a decrease in the expression to 0.25 and 0.7 folds for glnL and glnA, respectively. No notable changes were recorded in the amtB gene expression compared to that of control (Fig. 6B). The growth curve of both the mutated strains represented variations in growth with increased growth in LG37-OEglnK and decreased growth for LG37-ΔglnK strain when compared to the wild-LG37 strain (Fig. 6C). These results indicated that the GlnK plays a significant role in the NH4+ assimilation of LG37.
Based on the above results, we noted that the GlnK senses the NH4+ concentration and regulates glnL expression through signal transduction, and further, the GlnL promotes glnA and amtB in favor of NH4+assimilation. Besides, AmtB as an ammonium transporter, there was no significant expression among the 3 groups (Fig. 6B), which is similar to that of the transcriptome results of amtB between NH4+-N and Gln-N nitrogen groups. Hence, we analyzed the relative expression of AmtB in wild-LG37 at an increased NH4+ concentration, and the results exhibited no difference in their expression pattern even at the highest NH4+-N levels (Fig. 6D). The above data suggest that the GlnK plays a vital role in regulating GlnL and GlnA, and still, the AmtB were not as a specific positive factor for NH4+ transporter of LG37 in assimilation. Therefore, it has been ruled out that there might be other NH4+ transporters in the case of LG37.
Determination of the NH4+ transporterMnrA in LG37
Considering the relative expression of amtB indicated in Fig. 6B and D, we discovered that the AmtB is not specific in the NH4+ transport of LG37, but this finding was only based on our transcriptomic and mRNA expression data and not direct assessment. Thus, we further analyzed our transcriptome data for another transporter, where we noticed mnrA with a 2.93-fold change in expression. (Additional File 4). To verify whether MnrA plays a role in NH4+ assimilation, we detected the expression of mnrA of LG37 after culturing with different NH4+-N concentrations by using RT-qPCR. The expression of mnrA increased with an increase in NH4+-N concentrations (Fig. 7A). In the LG37-ΔglnK and LG37-OEmnrA mutant strains, the mnrA expression was increased by a factor of 1.67 and decreased to 0.35, respectively, compared to that of wild-LG37 strain (Fig. 7B). Similarly, we determined the growth of both strains showed increased growth pattern in LG37-OEmnrA, whereas LG37-ΔmnrA strain showed a decreased growth compared to that of wild-LG37 strain (Fig. 7C). These results demonstrated that MnrA is essential for the NH4+ assimilation process in LG37.
Further, to extend our studies on MnrA, we analyzed the expression of downstream functional genes glnA and glnB in the NH4+ assimilation pathway for both LG37-OEmnrA and LG37-ΔmnrA strains. The relative-expression levels of glnA (1.7 folds) and glnB (1.6 folds) were increased in the OEmnrA strains, whereas, in the ΔmnrA strain the expression of glnA and glnB were only 0.3 and 0.5 folds, respectively (Fig. 7D). These findings demonstrated that the MnrA was served as the transporter and play a significant role in the process of NH4+ assimilation in LG37.
{kind=link}
{kind=link}
{kind=link}