It is well known that the epigenetic regulation of imprinted genes is involved in many embryonic developmental processes, including cardiac development [17, 18].
At present, DNA methylation disorder is considered as one of the important factors in the occurrence and development of CHD[4]. However, the specific role of the imprinted gene methylation regulation on cardiac development remains unclear. In this study, we found for the first time the abnormal gDMR methylation changes of 8 imprinted genes (GRB10, MEST, PEG10, NAP1L5, INPP5F, PLAGL1, NESP and MEG3) in patients with CHD. The methylations of imprinted genes varied with the heterogeneity of CHDs. Six of them were associated with significantly higher risk of CHD except MEST and NAP1L5.
The etiology of CHD is complex, involving the interaction of environmental and genetic factors, leading to developmental phenotypes that regulate morbidity and severity. The potential relationship between alterations of genomic methylation and CHD is increasingly recognized. The dynamic changes of DNA methylation in cardiomyocyte-related gene sets during cardiac development suggest that DNA methylation modification is essential for the occurrence of cardiac diseases [3, 4]. One study has confirmed that there are significant differences in methylation of genes associated with muscle contraction and cardiomyopathy in CHD[4]. Genomic imprinting is a process of epigenetic modification, and loss of imprinting may lead to abnormal embryonic development [13]. DMR plays an important role in the establishment of gene imprinting by targeting imprinted gene. Deletion or abnormal methylation of DMR might result in the expression disorder of the imprinted gene cluster [19]. Specifically, we selected DMRs in the imprinting control region of imprinted genes studied, i.e., gDMRs, which are established during gametogenesis and play a key role in the imprinting of imprinted gene clusters [19, 20]. The small changes in gDMRs found in our study might be relevant in the etiology of CHD, which was supported by the stratified analysis. The relatively wide confidence interval in the risk analysis was due largely to the small sample size in this pilot study.
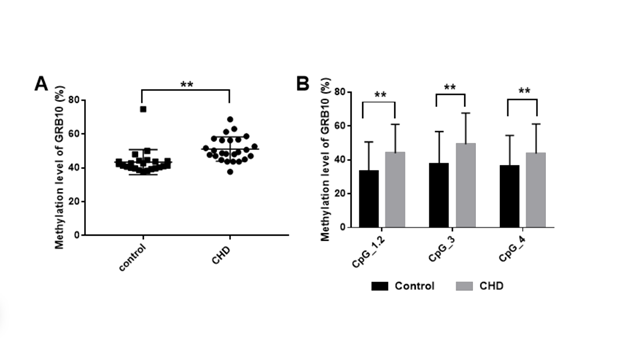
Some imprinted genes have key functions influencing the proper embryonic growth and development by forming "imprinted gene network", including GRB10 and MEST as the core molecules[13]. GRB10, as an important growth-limiting factor, has a wide range of effects on embryonic development. Maternal GRB10 knockout results in embryonic overgrowth, and the change of ICR of GRB10 gene results in obvious dysplasia [21, 22]. Previous studies have found an increase methylation of GRB10 in fetal samples of spontaneous abortion [23]. It has been found that the Ddc gene in mice, involved in the development of trabecular cardiomyocytes of the embryonic and neonatal heart, displays tight conserved linkage with the methylated GRB10 gene [24]. In the present study, we found that the gDMR methylation of GRB10 imprinted gene in CHD was increased except for valvar defects. It is hinted the methylation disorder of GRB10 may be involved in the development of heart through the interaction with Ddc. Hypermethylation of GRB10 might inhibit its expression and regulate the growth through insulin pathway such as IGF2, potentially leading to cardiac dysplasia. MEST is a maternal imprinted gene and widely expressed throughout the embryonic period [25]. Mest-knockout mice exhibited embryonic and placental growth retardation and postnatal growth inhibition, while loss of the imprinted gene resulted in postnatal weight gain and multiple organ hypertrophy [26, 27]. King et al. have found that Mest specifically expressed myocardial trabeculae in developing atria and ventricles of mice. Mest-knockout mice showed subtle changes of myocardial trabeculae, which displayed an increase in thickness and reduction in density of the compact myocardium, similar to that in human heart disease, which hinted MEST is closely related to heart disease [28]. The increased methylation of gDMR of MEST imprinted gene in CHD was also found in our data, especially in ASD and VSD. Potentially, MEST expression disorder via altered methylation of the gene might be involved in the occurrence of CHD by affecting the development of myocardial trabecula.
PEG10, a paternally expressed imprinted gene, is expressed in embryonic tissues and placenta, and participates in cell proliferation, differentiation and apoptosis [29, 30]. The Peg10-knockout mice exhibited early embryo death and placental defects [31]. A study has found that the expression of PEG10 in the heart was very low, indicating that trace PEG10 could maintain the normal development of the heart [29]. On the contrary, we found abnormal hypomethylation of PEG10 in CHD. Overexpression of PEG10 may be involved in the invasion and metastasis of malignant tumors, such as hepatocellular carcinoma and endometrial cancer etc., through the epithelial mesenchymal transition (EMT) [32, 33]. Therefore, the low methylation of PEG10 might cause abnormal elevation of expression, and induce abnormal migration of related cells through EMT pathway, potentially leading to abnormal cardiac development.
INPP5F encodes inositol 1,4,5-trisphosphate (InsP3) 5-phosphatase, which is an important functional endogenous regulator. It has been found that INPP5F regulated the size of cardiac myocytes and cardiac stress response, increased the hypertrophy and activation of the fetal gene program in the stress response in Inpp5f-knockout mice, and decreased the hypertrophic response in Inpp5f-overexpressed mice [34, 35]. INPP5F can be used as a negative feedback regulator of insulin signal and downregulation of INPP5F in diabetes mellitus has cardio protective effect [36]. Consistent with previous studies, we found that INPP5F methylation was reduced in a variety of CHDs and increased CHD risk, suggesting that INPP5F might be a protective factor for cardiac development.
MEG3 is a maternal-expressed imprinted gene that is expressed in many normal tissues. It inhibits the proliferation of tumor cells in vitro. It also interacts with tumor suppressor p53 to regulate the expression of p53 target genes [37]. Consistent with this effect, MEG3 has an impact on cardiac remodeling induced by pathological hypertrophy by regulating the binding of p53 to other gene promoters[38]. Other studies have shown that MEG3 participates in the development of myocardial fibrosis and prevents myocardial remodeling by regulating the production of MMP-2 by CFs in vitro and in vivo [39]. In consistency to previous studies, our results also found that methylation of MEG3 was reduced and associated with the increased risk of CHD.
We also detected hypomethylation of gDMRs of other 3 imprinted genes including NAP1L5, PLAGL1 and NESP in CHD. Stratified analysis revealed that different CHDs showed specific methylation patterns, such as hypomethylation of the 3 genes in ASD and VSD, hypomethylation of NAP1L5 and NESP in valvular malformation, hypomethylation of NESP in thoracic vascular malformation, and no significant change in ToF.
As a pilot study, it is subject to a couple of limitations. First, we studied a small and heterogeneous group of patients with CHD. A large sample size is warranted to identify the methylation characteristics and risk factors of imprinted genes in relation to the types of CHD. Second, the causal relationship between methylation alterations of imprinted genes and CHD remains to be explored in further experimental studies to examine their potential impact on RNA expression and CHD development on cellular and organ levels in animal models.
{kind=link}
{kind=link}