From the TCGA Breast Cancer dataset, gene expression data for 115 patients with triple-negative breast cancer and 112 normal adjacent tissues were retrieved. Fig. 1 demonstrates the study design, flow, and methodology used in the current study. In this cohort of 115 TNBC patients, favorable overall survival as per the Kaplan-Meier curves was significantly correlated with an earlier disease stage (P = 0.0012; Fig. 2).
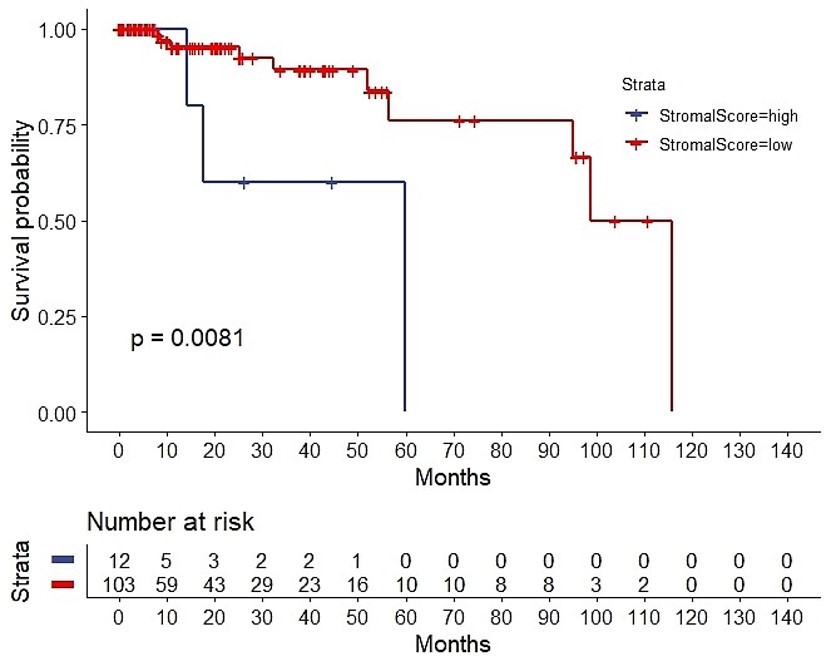
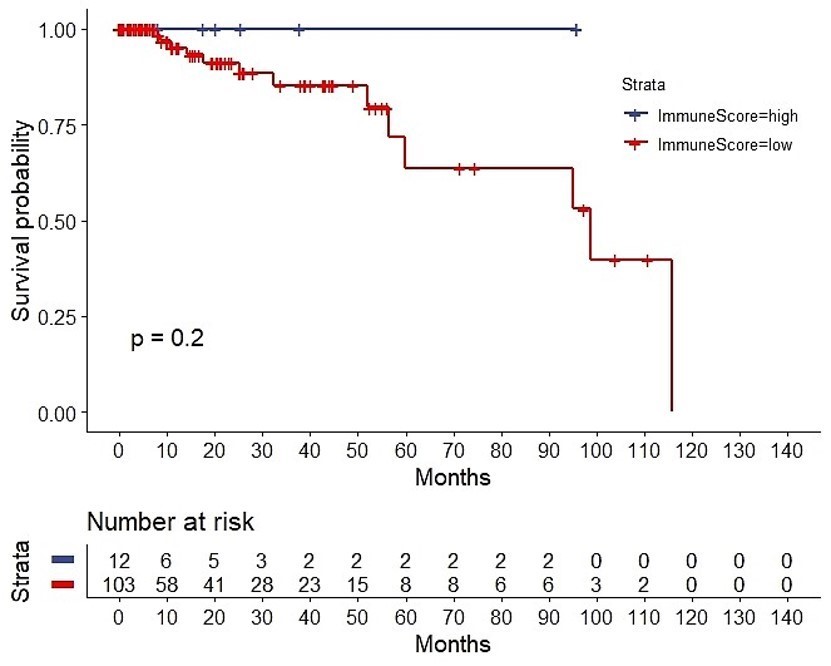
The ESTIMATE algorithm computationally estimates the fraction of stromal and immune cells in the TME. The SS was high (SS-high) in 10 patients, and the IS was high (IS-high) in the ten other patients. There were no significant correlations between the SS or IS and the cancer stage (Suppl. Fig. 1). However, as per the Kaplan-Meier analyses, SS-low patients showed a higher overall survival (OS) than that of SS-high patients (P = 0.0081; Suppl. Fig. 2), while IS-high patients showed a higher OS than that of IS-low patients (P = 0.2; too few cases in the IS-high; Suppl. Fig. 3). Of note, a strong correlation between both the SS and IS and the patients' overall prognosis was observed. Expectedly, when compared with IS-low/SS-high patients, IS-high/SS-low patients showed a better OS (P = 0.045) (Fig. 3). Cytoscape revealed the immune cell infiltration levels between samples grouped by IS-high/SS-low or IS-low/SS-high (Suppl. Fig. 4). Interestingly, grouped linear regression showed a statistically significant increase in M2 macrophages in TNBC patients with the IS-high/SS-low phenotype. In line with these results, in the tumor microenvironment (TME), a higher proportion of M2 was also found in IS-low/SS-high patients, as estimated by CYBERSORTx (P < 0.001) (Suppl. Fig. 5). Additionally, significantly more CD8+ cytotoxic T-cells, memory B cells (P = 0.0304), activated CD4+ memory T cells (P = 0.0056), follicular helper T cells (P = 0.0044), and activated NK cells (P = 0.0511) were also observed in IS-high/SS-low patients (14% vs. 5.3%, p = 0.0143) (Table 1). On the other hand, more resting NK cells were observed in IS-low/SS-high patients (P = 0.0108).
Moreover, DEG analysis showed 651 DEGs (284 upregulated, and 367 downregulated) in IS-high/SS-low patients, and 370 DEGs (187 upregulated, and 183 downregulated) in IS-low/SS-high patients (Suppl. Fig. 6). Heatmaps and volcano plots of the DEGs in the context of these two groups are shown in Fig. 4A-D. KEGG pathway enrichment analysis showed that the overexpressed DEGs from both phenotypes were enriched in the IL-17 signaling pathway and viral protein interaction cytokine and cytokine receptor genes. Notably, the overexpressed DEGs in the context of the SS-low/IS-high phenotype were also enriched in other two cytokine-related pathways: cytokine-cytokine receptor interactions pathway and chemokine signaling pathway (Fig. 4E).
Using X2K, we also inferred the transcription factors associated with the two immune phenotypes. In SS-high/IS-low TNBC patients, the inferred TFs were PPARG, HNF4A (also known as farnesoid X receptor, FXR), NR1H4, NR0B2, and MLXIPL, whereas the TF signature of SS-low/IS-high TNBC patients was composed of the PPARG, CEBPA, and MLXIPL TFs (Fig. 5). Of note, the OncoScore analysis did not perform as well in the search for the TF signatures (Suppl. Table 1). Additionally, we performed a manual search on PubMed and discovered nine TF genes linked to IL-17-mediated signaling, including PPARG, CEBPA, MEOX1, KLF15, CD36, ZNF750, EZH2, HNF4A, and NR0B2 (Table 2).
Importantly, GSVA suggested the involvement of additional pathways, including the JAK-STAT signaling, T cell receptor signaling, B cell receptor signaling, cytokine-cytokine receptor interaction, TGF-β signaling, and PPAR signaling pathways (Fig. 6; the upper panel shows the GSVA of the 370 DEGs of the SS-high/IS-low subgroup and the lower panel shows the GSVA of the 651 DEGs of the SS-low/IS-high subgroup).
Validation of the results in the cBioPortal
IL-17 is a proinflammatory cytokine that signals mainly via the TRAF3 Interacting protein 2 (TRAF3IP2), as reported previously [27, 28]. TRAF3IP2 is an inflammatory mediator and upstream regulator of several crucial transcription factors, such as AP-1 and NF-κB [28]. Act1, an essential component in IL-17 signaling complex, is encoded by the gene TRAF3IP2. Finally, we used the Oncoprint from cBioPortal to validate the frequency of IL-17 genes in TNBC. We discovered that 13% of ER-negative and HER2-FISH unamplified breast cancers harbored IL17RA deep deletions and 25% harbored TRAF3IP2 amplifications (Suppl. Fig 7).
Interestingly, we also discovered using the cBioPortal platform that aberrations in the nine TF genes mentioned above are associated with a worse prognosis, as per the relapse-free survival of 2,377 patients (log-rank, P = 0.00007) and the overall survival of a larger group of 4,819 patients (log-rank, P = 0.001697) (Fig. 7).
{kind=link}
{kind=link}