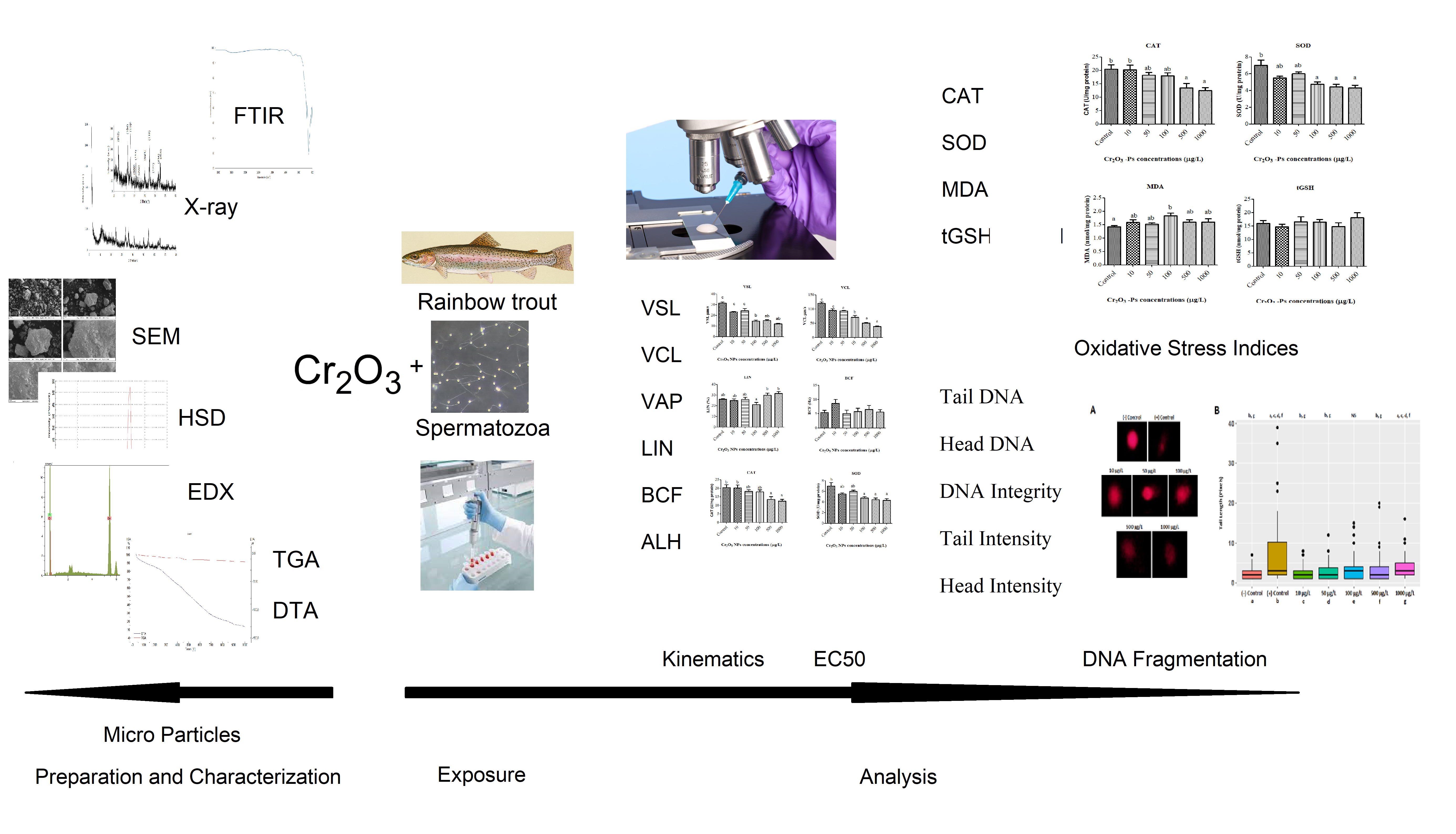
This study investigated the cytotoxic effects of different doses (10, 50, 100, 500, and 1000 µ/L) Cr2O3-Ps on spermatozoa of rainbow trout (Oncorhynchus mykiss) in vitro conditions. For this target, the cytotoxic effects were analyzed the functional deformations in the velocities, movement styles, enzymatic activities, and DNA damages of spermatozoa, the end of the study was determined effective concentrations of Cr2O3-Ps.
In this study, Cr2O3-Ps caused changes in the physical and biochemical functions in spermatozoa of rainbow trout. In light of the literature for functional deformations, there are same results which are parallel with our data. For example, lower doses than 100 µg/mL dose of Cr2O3-Ps showed minimum toxicity on viability or apoptosis of two human lung cell lines, BEAS-2B and A549 (Chusuei et al. 2013). Another hand, high doses such as 300 cm2/mL of Cr2O3-Ps had a negative on the function and mechanism of epithelial cells, monocytic/macrophage cells, human erythrocytes, and combined culture (Cho et al. 2013). However, it was observed morphological damages and partially disintegration in body of Daphnia similis (Tavares et al. 2014). However, especially the functional negative effects were shown on the enzyme activities and DNA fragmentations of spermatozoa with dose-dependent manner of Cr2O3-Ps in our study. Similarly, it was determined the negative effects such as significant growth inhibitory and cell membrane damage and oxidative stress responses in ROS on Escherichia coli after exposure of Cr2O3-Ps (Kaweeteerawat et al. 2015). The exposure to Cr2O3-Ps on green alga showed cytotoxic effects such as inhabitation growth, lowered chlorophyll content, increasing ROS levels (Costa et al. 2015). Other hand, it was found the negative biological functions such as longevity, reproduction, and growth parameters of Daphnia magna after Cr2O3-Ps exposure (Puerari et al. 2016) and the diversity of bacterial communities and inhibited the enzyme activity analysis (Wang et al. 2017).
Nano or micro particles have also harmful effects on the functional structures of cells such as reduced cell viability and inhibited proliferation (Agrawal and Kango 2019), reduction of spermatozoa velocities (Özgür et al. 2018a) with DNA damage and enzyme activities (Préaubert et al. 2018; Agrawal and Kango 2019; Barkhade et al. 2019; Nikolovski et al. 2019; Santonastaso et al. 2019). In our results, the velocities (VSL, VCL, and VAP) and the movement styles (ALH and LIN) of spermatozoa were negative affected by dose-dependent manner of Cr2O3-Ps, and these results were parallel with data of above literatures.
The cytotoxic effects of Cr2O3-Ps were observed changes in enzymatic activities in spermatozoa. Biochemical parameters of spermatozoa such as CAT, SOD, MDA and tGSH as oxidative stress indices were measured after Cr2O3-Ps exposure. SOD catalyzes the conversion of superoxide (O2−) radical to H2O2, while CAT carries out the conversion of H2O2 to water and oxygen. Therefore, the SOD-CAT system provides the first defense against oxygen toxicity (Afifi et al. 2016). According to our results, there was a significant decrease in the levels of CAT and SOD with increasing concentrations of Cr2O3-Ps. This decrease in both SOD and CAT levels may have been caused by the excessive ROS production induced by Cr2O3-Ps. The same results were reported by Afifi et al. (Afifi et al. 2016). They investigated the toxicity effect of Ag-NPs on Oreochromis niloticus and Tilapia zillii. While 2 mg/L Ag-NPs did not lead to any significant change in the SOD and CAT levels, dose of 4 mg/L showed a significant reduction in the levels. Similarly, our previous study reported that SOD and CAT levels were significantly decreased after exposure to 100 mg/L of Fe3O4 NPs (Özgür et al. 2018b). In our study, While MDA level significantly increased compared control group, tGSH levels did not change. MDA is an important biomarker to determine lipid peroxidation. This increase in MDA level after exposure to the Cr2O3-Ps could be due to the depletion of the antioxidant system, which is consistent with the aforementioned results. Also, Cr ions produced from Cr2O3-Ps are incriminated from increased lipid peroxidation (Sinha et al. 2005). Additionally, our outcomes were enforced by the results of Adebayo et al. (Adebayo et al. 2018). Other hand, it is well known that GSH protects the biological systems from oxidative stress. Mechanisms of change in the tGSH level may be different. For instance, excessive ROS production may have affected tGSH levels. Alternatively, the Cr ions may have shown inhibition on GSH-synthesizing enzymes. Maybe both are involved.
Investigating the effects of UV radiation and hydrogen peroxide (used as oxidative agents) on rainbow trout, Oncorhynchus mykiss, spermatozoa, Dietrich et al. (2005) found a decrease in sperm motility and DNA integration after a long duration of UV. From our research, H2O2 effect on a spermatozoa cell has been observed as (+) Control from Fig. 2A. Dose-dependent reductions in sperm motility and fertilizing features were significantly changed after spermatozoa were exposure to H2O2. Exposed of rainbow trout spermatozoa to some elements such as mercury and cadmium occurs an increase in DNA damage measured via Comet assay with brutal results in terms of sperm motility and hatching rate (Dietrich et al. 2005). Unlike oocytes, fish spermatozoa have been known not to have efficient evolutional defense mechanisms like DNA repairing against environmental pollutants even though having extremely condensed genetic material, DNA (Aitken et al. 2004). Furthermore, after exposure to physical/chemical stresses, spermatozoa have exhibited extreme sensitivity to oxidative stress, which is responsible for DNA fragmentation, because of not having enough antioxidant defense mechanism and extreme content in unsaturated fatty acids (Labbe et al. 1995; Cabrita et al. 2010; Li et al. 2010a, b; Linhartova et al. 2013).
{kind=link}