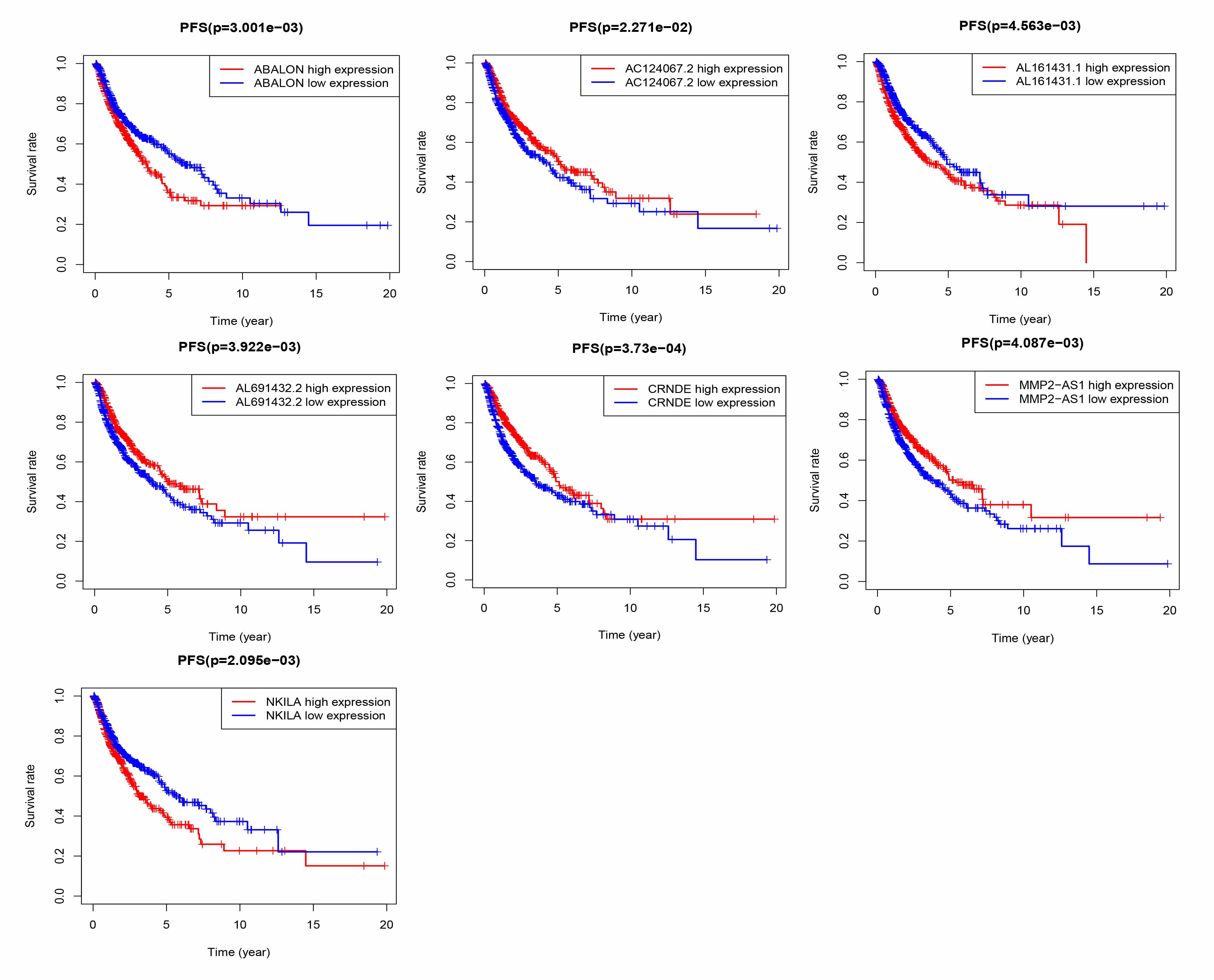
In our study, we identified and independently verified prognostic signals based on seven ATG lncRNAs (AL691432.2, MMP2-AS1, AC124067.2, CRNDE, ABALON, AL161431.1, and NKILA). GSEA results showed that risk signals were significantly correlated with immune cell function and immune response. This finding led us to further study the relationship between autophagy and immune infiltration.
lncRNAs are a type of noncoding RNA with a length of more than 200 nucleotides. They interact with proteins, RNA, and DNA to regulate gene expression before and after transcription. Studies have found that lncRNAs are widely involved in human life activities such as cell development, differentiation, proliferation, apoptosis, migration, invasion, and metastasis, which are closely related to the formation, progression, and metastasis of malignant tumors. In addition, lncRNA is also involved in chemotherapy resistance and targeted therapy.
A study of endometrial cancer found that lncRNA AL161431.1 targets miR-1252-5p, which leads to the upregulation of MAPK signals and regulates cancer cell apoptosis and autophagy cell death [13]. NKILA is a lncRNA that interacts with NF-κB; it regulates the sensitivity of T cells to T lymphocyte death (AICD) by inhibiting NF-κB activity. NKILA can directly or indirectly inhibit IκBα phosphorylation and NF-κB activation in breast and liver cancers [14, 15]. In breast cancer studies, inflammatory cytokines were found to upregulate NKILA expression through the NF-κB pathway, which regulates breast cancer metastasis and inhibits angiogenesis in HUVECs [14, 16]. In nasopharyngeal carcinoma (NPC), NKILA was also found to inhibit NPC metastasis through the NF-κB pathway [17]. P65 in laryngeal cancer tissues can also reportedly positively regulate NKILA expression; however, NKILA can inhibit the transport of P65, thus reducing the resistance of cells to laryngeal cancer X-ray radiation [18]. We found that NKILA reduces MMP14 expression by mediating IκBα phosphorylation and NF-κB translocation to the nucleus, thereby impairing the migration and invasion of ESCC cells [19]. Lung cancer studies have found that NKILA is associated with the survival prognosis of LUAD NKILA expression is regulated by the classic TGF-β signaling pathway, which subsequently inhibits the migration and invasion of NSCLC cells by interfering with the NF-κB/Snail signaling pathway in NSCLC cells [20]. A similar function was observed in tongue squamous cell carcinoma [21].
Colorectal neoplasia differentially expressed (CRNDE) was first observed in colorectal adenomas and colorectal cancers [22]. CRNDE is elevated in a variety of cancers, including colorectal cancer (CRC) [23, 24], glioma [25], hepatocellular carcinoma [26], and lung cancer [27], and there is increasing evidence that it plays a role in regulating cancer cell proliferation, migration, invasion, and apoptosis. Downregulation of CRNDE reportedly results in the inhibition of CRC cell proliferation and induces cell apoptosis [24]. Ellis et al. found that CRNDE is related to the regulation of aerobic glycolysis, or the Warburg effect, in cancer cells [28]. In lung cancer studies [29], LUAD tissue and CRNDE were significantly associated with poor differentiation, TNM stage, lymph node metastasis, radiotherapy response, and shorter OS time. Liu et al. found that CRNDE promotes NSCLC cell proliferation and growth by activating PI3K/AKT signal transduction [30].
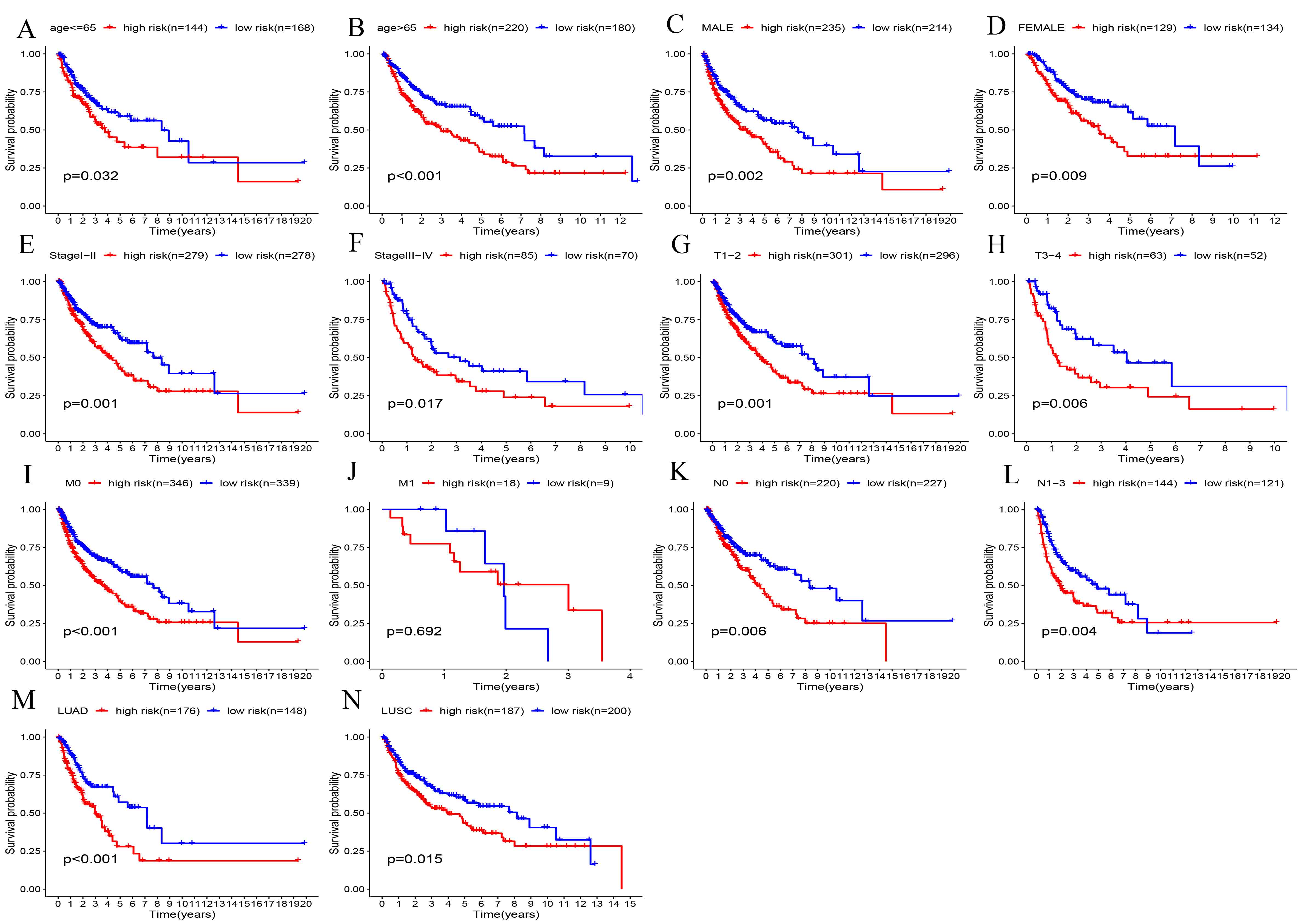
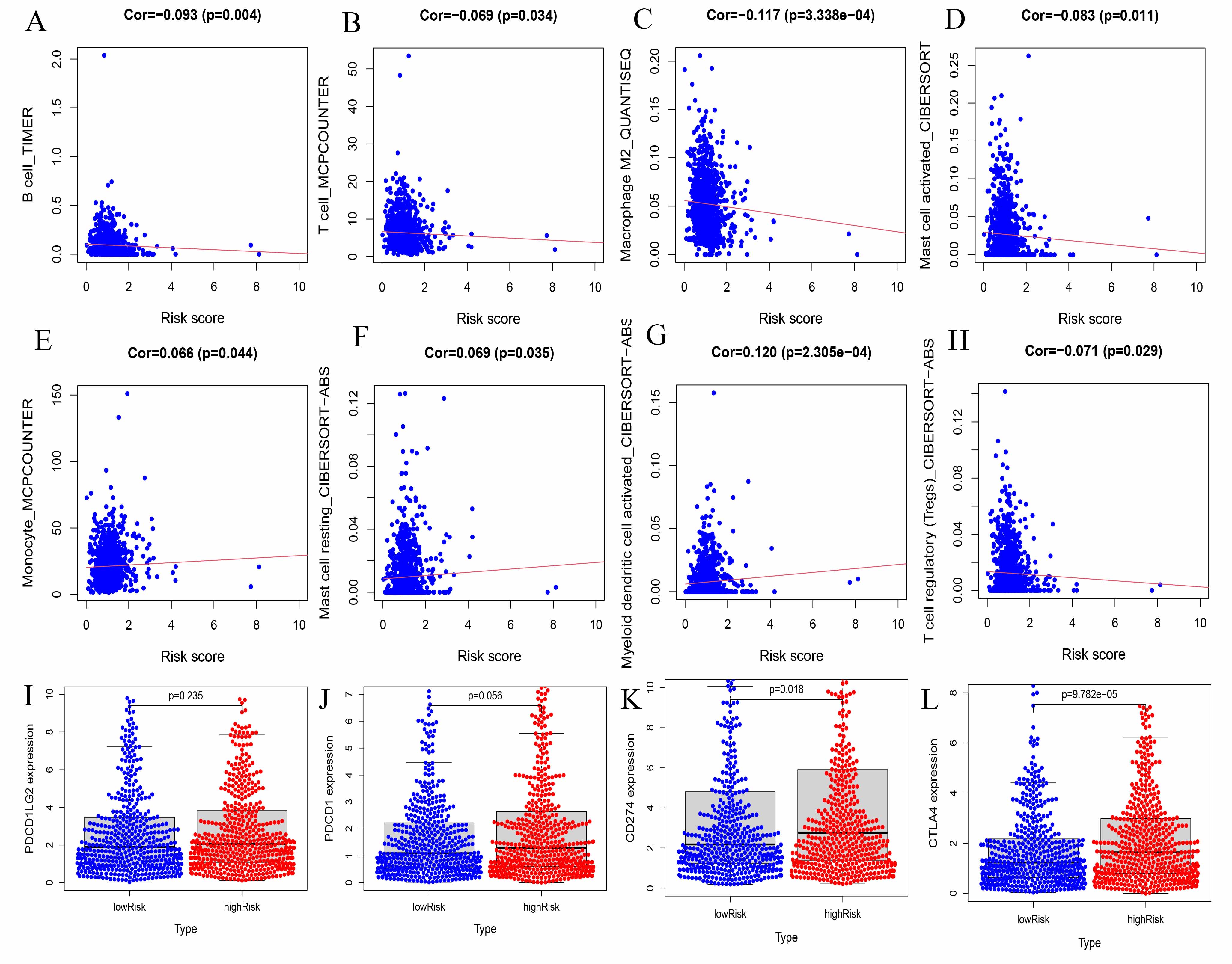
As tumor-infiltrating lymphocytes influence the immunotherapy response as well as the clinical consequences of colorectal, gastric, lung, and breast cancers, their role in various cancer-related processes is increasingly recognized. In this study, we also used gene expression profiles to explore the involvement of TILs in NSCLC patients. In our ATG risk model, the high-risk and low-risk groups, showed a different immune landscape. Specifically, we found that the distribution of B cells, T cells, macrophages, DCs, mast cells, and monocytes in our risk model was different and correlated with risk (P < 0.05).
Increasing numbers of studies are using DCs to treat cancer. DCs have strong antigen presentation ability, which may overcome tumor tolerance and induce antitumor immunity [31]. In NSCLC patients, DCs upregulate the expression of the coinhibitory molecule B7-H3 and inhibit T cell activity [32]. Lung tumor cells secrete substances that lead to monocyte-induced DC differentiation disorders, maturation abnormalities, and phenotypic defects. Tumor-induced DC deficiency leads to an insufficient ability to recognize and present tumor antigens and tumor immune escape [33].
Previous studies have shown that T cells are the main immune-infiltrating cells in NSCLC [34]; among these cells, CD4+ T cells are the most abundant T cell population (26%), followed by CD8+ T cells (22%) [35]. Many studies have shown that low levels of CD8+ T cell infiltration in tumor lesions are associated with poor prognosis [36]. As a key factor in the tumor microenvironment, B cells and neutrophils play an important regulatory role in tumor progression. However, these cells have both antitumor and pro-tumor activities [37–39].
The same macrophages play different roles in the tumor microenvironment. In LUAD, there were higher immune scores, more memory B cells, and more M0 macrophages in the early stage than in the late stage. The abundance of M0 macrophage infiltration was significantly correlated with TNM stage and survival rate [40].
In LUAD, mast cells are associated with tumor angiogenesis and poor prognosis [41, 42]. In this study, a predictive model of autophagy-related lncRNAs was established and validated by univariate and multivariate Cox regression analyses. The value of our K-M survival curve was P = 3.325e − 05, indicating that our prediction model was closely related to the survival outcomes of patients with lung cancer. In addition, our prognostic model was superior to the other prognostic markers. Finally, we used GSEA to detect the biological functions of the prediction model. These results strongly demonstrate that these lncRNAs are involved in tumor progression and are associated with immune infiltration.
Our study has several limitations. First, our findings need to be further validated in other independent cohorts to determine the stability of autophagy-related lncRNA prognostic markers. Second, our study was based on 712 patients from the publicly available TCGA database. Among them, the samples of NSCLC patients with early cancer (n = 557) were significantly larger than those of patients with advanced cancer (n = 155), which may have skewed our results; therefore, further analysis with a larger sample size is needed. Finally, further studies on biochemical experiments, such as immunohistochemistry, quantitative real-time polymerase chain reaction, flow cytometry, and clinical data analysis, are needed to confirm our findings.
{kind=link}
{kind=link}
{kind=link}
{kind=link}