Mosquito strains
Two strains were used for the experiments; the Kisumu strain of An. gambiae s.s, colonized over 40 years ago, from the area of Kisumu, Kenya, East Africa and the 17-year-old Mopti strain of An. coluzzii, colonized in 2003 by the Lanzaro Laboratory (UC Davis) from the village of NʼGabacoro droit near Bamako, Mali, West Africa. Natural West-African populations of the sibling species have recently introgressed with one another, resulting in the possible selective introgression of important pesticide resistance and ammonia detoxification loci (28,29). Therefore, in this study, we favoured older strains colonized before the intensification of chemical control with as little evolutionary history of such recent anthropomorphic selection pressures and associated introgression (27,29,30).
Strain maintenance
The strains were maintained by the Tripet group in dedicated insectaries at the Centre of Applied Entomology and Parasitology (CAEP), Keele University, UK. Mosquitoes were maintained at 25 ± 2 °C, a relative humidity of 70 ± 5%, with a 12-h light/dark photocycle. Larvae were fed an optimized diet of groundfish food (Tetramin, Tetra, Melle, Germany) at a rearing density of 200 larvae/litre by manual counting (31). Pupae were transferred to 5l plastic tub cages (20.5cm height × 20cm diameter), covered with netting for adult emergence. Cages had sleeved opening for easy management of mosquitoes and accessories. Approximately 600–800 adults were held in a cage, sugar was provided via a paper towel soaked in 10% glucose solution, and water via a soaked cotton pad in an upturned bowl placed on the cage netting. Female adult mosquitoes were fed with horse blood using an artificial feeding membrane (Hemotek feeding membrane system, Discovery workshops, Blackburn, UK). Styrofoam cups (egg cups) containing filter paper and water were placed in the cages 4 days after blood-feeding, to collect eggs. Following the removal of the egg cups, the cages were washed thoroughly and sterilised with bleach. Mouth aspirators were used to transfer adults from one container to another when necessary.
Experiment 1: Effects of ammonia, feed regimes and mineralization on the developmental success of An. gambiae s.s. and An. coluzzii.
Experimental design
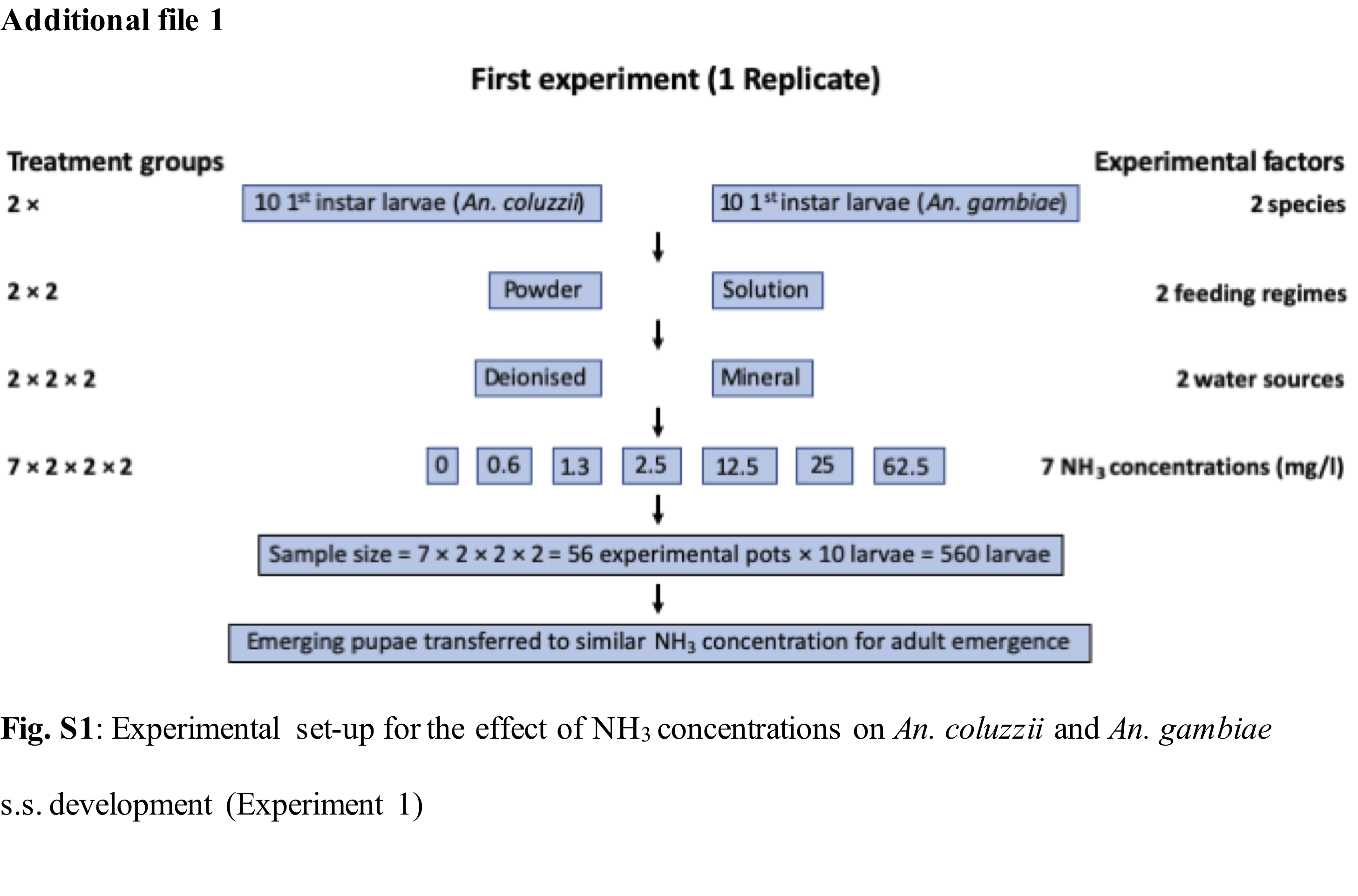
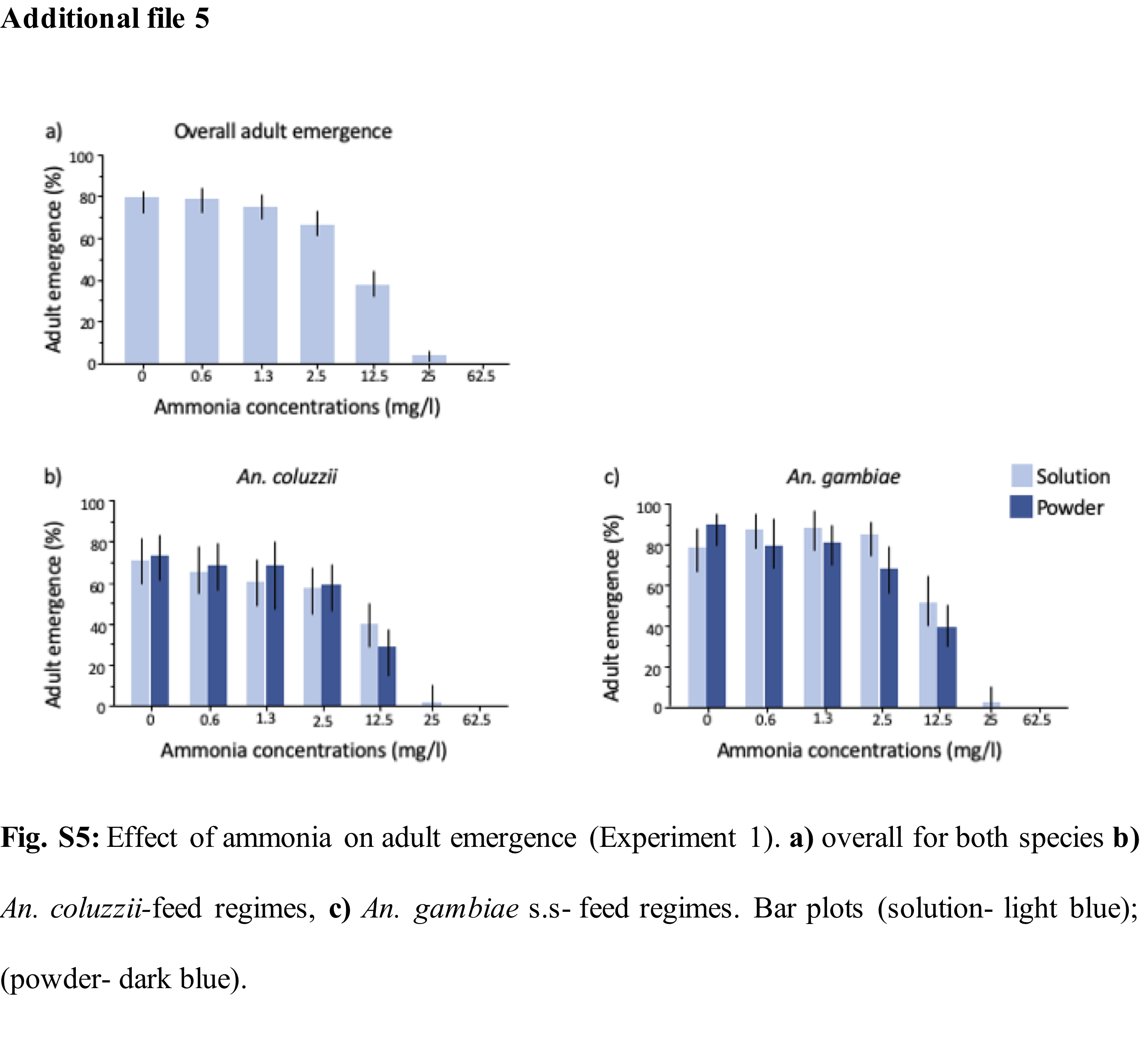
The experimental design consisted of 2 species, 2 feeding regimes, 2 water types and 7 ammonia concentrations resulting in 2 x 2 x 2 x 7 treatment groups (Additional file 1: Fig. S1). First instar larvae of An. coluzzii and then, An. gambiae s.s. were distributed using a 3ml plastic pipette and kept in groups of 10 larvae in white styrofoam cups containing 300ml of water (at 5cm depth) with variable ammonia concentrations. Three replicates were made resulting in a total of 1680 (560 × 3) larvae used in the study. To keep the ammonia and mineralization constant and unaffected by the accumulation of waste products, larvae from every group were transferred daily into new containers containing fresh water of their respective treatment group quality. At pupation, pupae were transferred to a netted styrofoam cup containing 100ml of the same water quality but with no ammonia. The position of experimental pots was completely randomized, and experimental cups labelled with codes to avoid bias due to variation in the insectary environment.
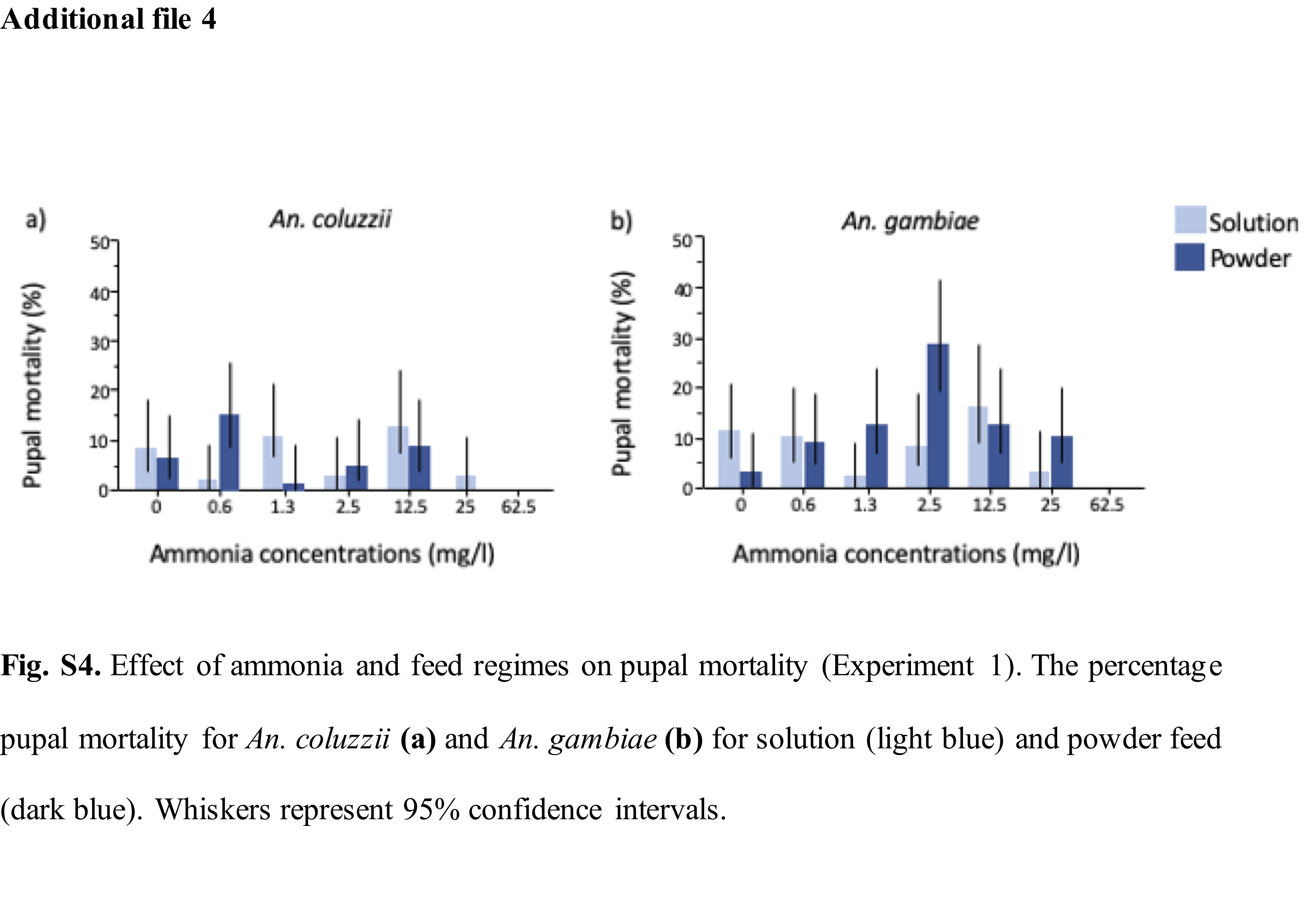
Two standard feeding regimes were used: solution or powder feed. On day 1, larvae from both feeding regime groups received a 0.1ml single drop of Liquifry liquid fish food (Interpret Ltd, Surrey, UK). From day two, half the cups were fed on a powder feeding regime that consisted of daily rations of TetraMin Baby powder fish food (Tetra GmbH, Melle, Germany). The rations increased over time and were dropped on the water surface using a micro-spatula where it spread on the water surface (2mg on days 2–3, 4mg on day 4, and 10 mg on day 5 until pupation). The solution feeding regime consisted of the same food quantity dissolved in deionized water (0.1ml of 1g/50ml of TetraMin Baby on days 2–3, 0.2ml of 1g/50ml of TetraMin Baby on day 4, and 0.5ml of 1g/50ml of TetraMin Baby on day 5 until pupation) and injected into the larval pot using a pipette.
Mosquito larvae were reared in two water types with different levels of mineralization: (i) deionised water which was sourced from a reverse osmosis unit (PURELAB Prima, Wycombe, United Kingdom) installed in the laboratory. The water quality specifications of treated deionised water were: TDS (27.55 ± 2mg/l), salinity (18.48 ±1ppm), conductivity (39.54 ± 2µS), total organic carbon < 0.1ppm, bacteria < 5 CFU/ml, 98% rejection of inorganics, > 99% rejection of organics, > 99% rejection of particles. (ii) mineral water containing natural minerals formed through geological processes and sourced in 5l bottled from a local supplier. Water quality specification for mineral water were: TDS (112.21 ± 2mg/l), salinity (75.78 ±1ppm), conductivity (160.40 ± 2µS) This water contained the following minerals per litre: calcium (11mg), magnesium (3.5mg), potassium (2.5mg), sodium (10mg), bicarbonate (25mg), sulphate (11mg), nitrate (15mg), chloride (14mg), dry residue at 180 °C (85mg) and its pH was 6.2.
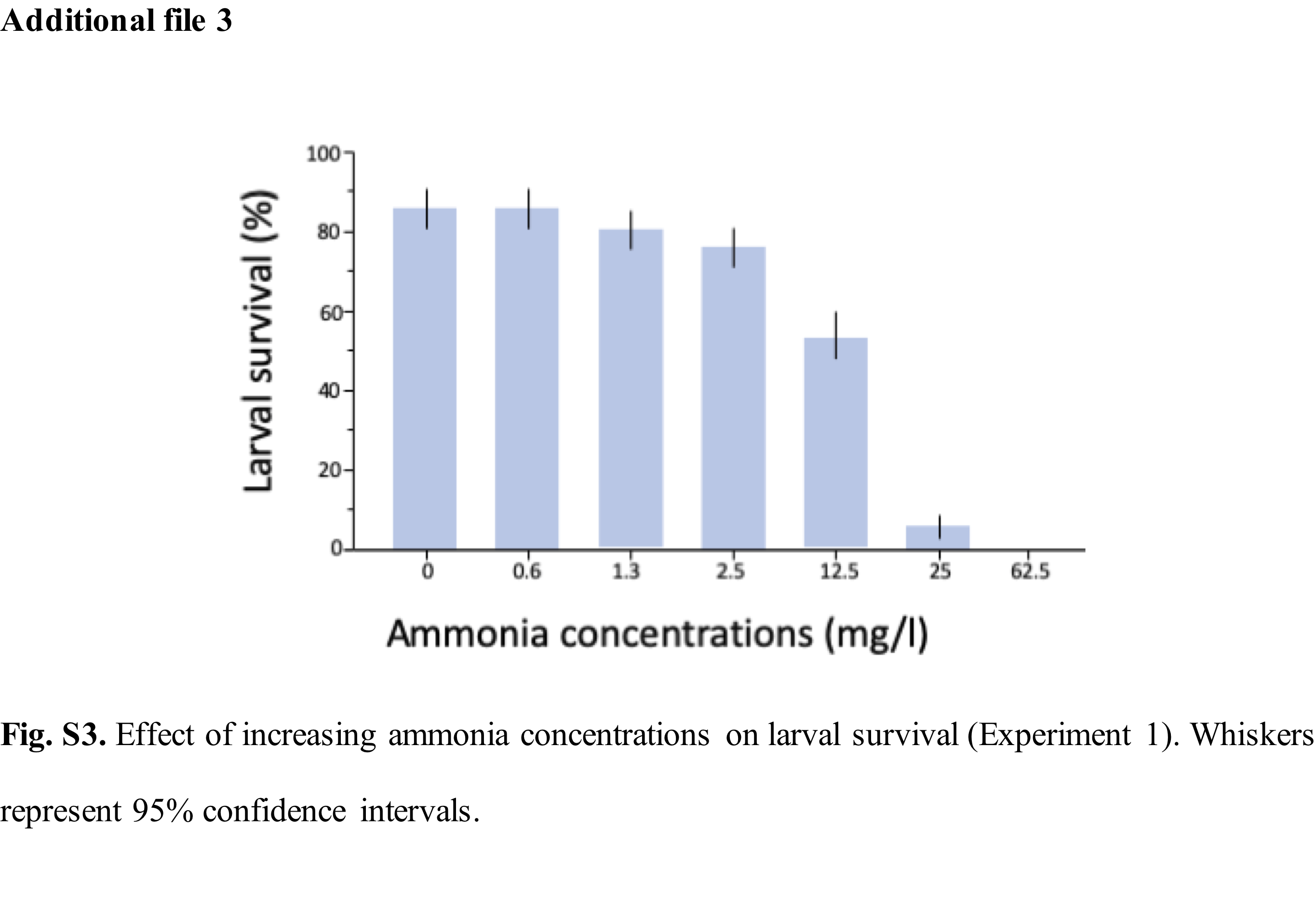
For each water type and feeding regime, mosquito larvae were reared in 7 ammonia (NH3) concentrations from 0mg/l (control); 0.6mg/l; 1.3mg/l; 2.5mg/l; 12.5mg/l, 25mg/l and 62.5mg/l, informed by published reports of the natural levels of ammonia in rice field flood water which are equivalent or higher than 0.5mg/l and the toxicity level of unionised ammonia to freshwater aquatic organisms at concentrations above 0.2mg/l (22–25,32). A stock solution of ammonium hydroxide equivalent to ~ 28-30% NH3 at 14.8M (Sigma-Aldrich, Missouri, USA) was diluted to 1% NH3 (~2.5g/l) by adding a proportionate volume of water. Further dilutions were then made using the C1V1 = C2V2 dilution formula.
Developmental success parameters
Depending on the life-cycle stage of the mosquitoes, the following data were observed and recorded: (i) larval survival: determined as the percentage of larvae that developed into pupae from the total number of larvae for each treatment; (ii) pupal survival: determined as the percentage of mosquitoes that emerged as adults from those that pupated in each treatment. (iii) pupal mortality: determined as the percentage of mosquitoes that died at the pupal stage from the total number of mosquitoes per treatment; (iv) adult emergence: determined as the percentage of mosquitoes that emerged as adults from the total number of larvae in each treatment; (v) development time: determined as the number of days from placement of first instar larvae in treatment cups until adult emergence; and (vi) wing-length: following emergence, adult mosquitoes were sexed and stored in 75% ethanol. One wing of all emerged adults was measured from the distal end of the allula to the apical margin (radius veins), excluding the fringe scale using a binocular microscope. A stage micrometre of 1mm ruler length (Graticules Ltd, Kent, UK) was used for calibration on 2.5 magnification on a scale of 1 microscope unit = 0.04mm) (33).
Experiment 2 - Developmental success and phenotypic quality of An. gambiae s.s. and An. coluzzii in contrasted microcosms.
Experimental design
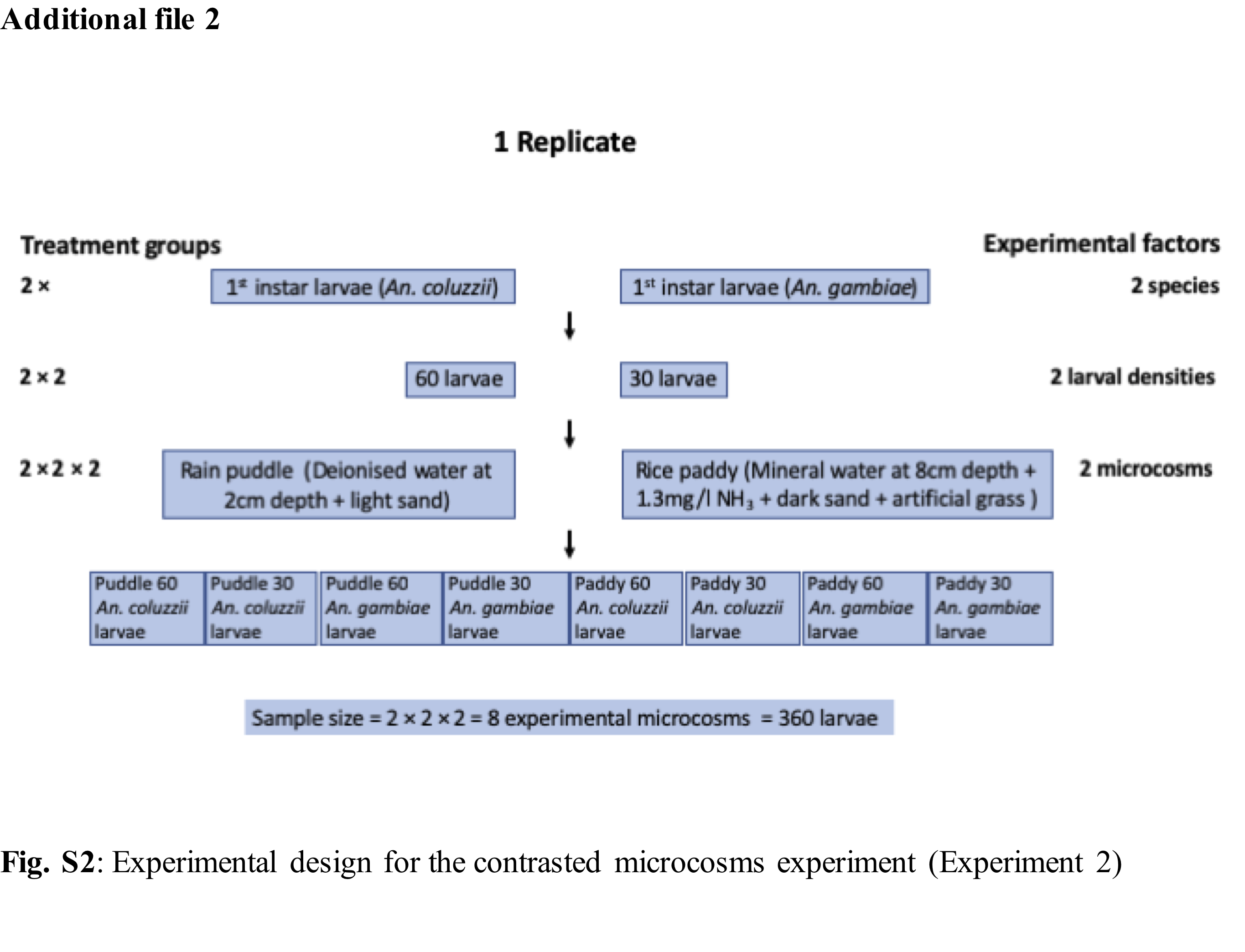
To investigate the developmental responses of the sibling species in contrasted larval habitats, first instar larvae of An. coluzzii and An. gambiae s.s. were reared in rain puddle and rice paddy simulated microcosms. Rain puddle (An. gambiae s.s. preferred) larval microcosms were simulated with transparent aquarium-like containers of 19.5cm length, 16.5cm height and 12.5cm width, containing deionised water 4cm deep and 400ml of wet inert light brown sand (UNIPAC aqua gravel, Northampton, United Kingdom) of 2cm depth (Fig. 1a,b). Rice paddy (An. coluzzii preferred) larval microcosm was simulated using a similar container, containing mineral water at 8cm depth, 1.3mg/l of ammonia, 400ml of inert dark sand 2cm deep (sourced as before), and a patch of intermediate density plastic grass (to imitate the rice field), 11.3cm length × 6.2cm width (patches were 5cm apart) (Fig 1c,d).
Each replicate consisted of two larval densities (30 and 60 larvae) of first instar larvae of both species placed in the two types of microcosmic larval habitats (2 species × 2 densities × 2 habitats = 8 microcosms), resulting to a total of 720 sampled mosquito larvae for two replicates. The total of 16 microcosms was randomly arranged and rearranged daily from day 5, to avoid confounding effects. To prevent ammonia build-up, on day 5 and afterwards, every other day until the completion of the assay, water was pumped out from the microcosm using a low-pressure pump, down to a minimal level to avoid picking up larvae and replaced with fresh water as per larval habitat type. Microcosms were covered with mosquito netting material to contain the emerging mosquito adults and entry of external debris. Coloured tapes were used to differentiate the habitat type and species, the microcosms arrangement in the insectary was completely randomised (Additional file 2: Fig. S2). All larvae were fed daily following a standardised feeding regime consisting of 0.1ml of Liquifry liquid fish food (Interpret Ltd, Surrey, UK) on day 1. From day 2 until pupation, finely ground Tetramin flakes (Tetra GmbH, Melle, Germany) was mixed with deionised water and injected into the microcosm in the following proportions: day two to three - 0.25ml of 0.2g/10ml solution of ground Tetramin flakes; day 4 - 0.5ml of 0.16g/10ml solution of ground Tetramin flakes; day 5 until pupation - 1ml of 0.75g/50ml of ground Tetramin flakes.
Developmental success parameters
Adult emergence was recorded as the number of adults that emerged from the total number of larva placed in a microcosm. Development time was recorded as the duration between day 1 of the experiment when first instar larvae were placed in microcosm and the day of adult emergence. Adults that emerged from the microcosms were collected using a mouth aspirator, sexed and stored in 75% ethanol for subsequent measurement of wing-length as described in first experimental design.
Physicochemical water parameters
Levels of nitrates, ammonia, pH, general hardness and carbonate hardness were measured using API aquarium test kits (Aquarium Pharmaceuticals, Mars Fishcare, Chalfont, USA). Readings were taken 10 days after the experimental set-up for the first experiment (Table 1) and on days 1 and 10 for the second experiment (Table 2)
Statistical analysis
All data collected were analysed using the software JMP 14 (SAS Institute, Inc., Cary, North Carolina, USA). All data were checked for deviations from normality and heterogeneity, and analyses were conducted using parametric and non-parametric methods as appropriate. Replicate effects were tested but are only reported when significant. Interactions between independent variables were tested using a step-wise approach whereby only those significant were retained in the final models. Following logistic regressions on proportions of larvae, pupae and adults, likelihood odds ratios were used for post-hoc pairwise group comparisons. Body size, a continuous variable, was analysed through general linear models followed by Tukey’s HSD post-hoc pairwise comparisons. Finally, developmental times (day of emergence) were analysed by Cox Proportional-Hazard models with likelihood odds ratios for post-hoc pairwise comparisons.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}