Mutual interaction, signalling and cooperation between the ECM and the cell, and, moreover, the principle of feedback in these relationships are key factors determining the fate of the cell [5, 25]. In the present research, we wanted to clarify whether a change in the basic physicochemical parameters of the surface (mimic ECM) will change the behaviour of the cell, and especially whether it will reduce its carcinogenic potential. However, based on previous studies, we have shown that modification of the surface ECM changes the cell's demand for functional molecules so that the surface effect also depends to a large extent on the chemical compounds available to the cell, mainly proteins [19, 26]. Therefore, an attempt was made to force the cell to change its behaviour (weakening its invasiveness) by physicochemical modification of its niche together with supplying its environment with a wide spectrum of potentially useful proteins. For the first time, it has been shown that the use of mechanical stimulation with the addition of a protein cocktail can activate transmembrane signalling mechanisms and consequently decrease the proliferation of HepG2 liver cancer cells and reduce their carcinogenic potential.
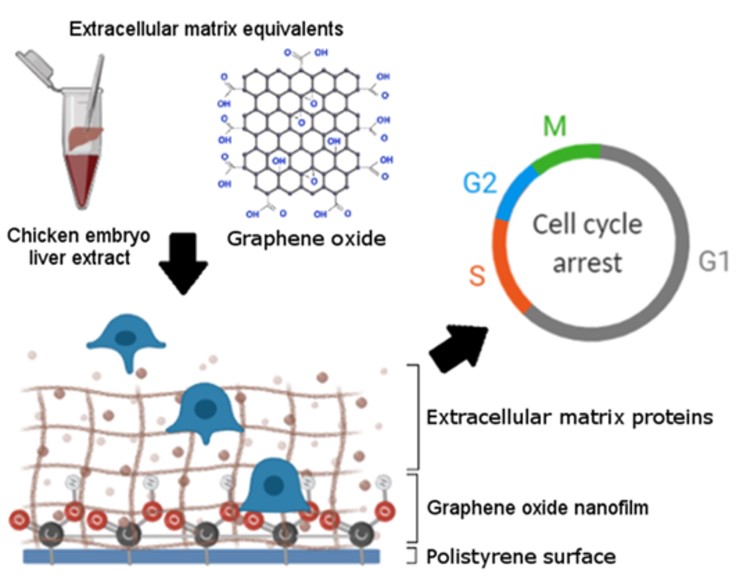
In the present study, the surface of the culture plate was modified by a thin layer of GO which formed a nanofilm that could act on receptors located on the cell membrane but was not internalised by cells [27]. The newly created surface (nfGO) was a source of mechanical and chemical signal modification. The reduction of stiffness (2.3 GPa) and the increase in roughness by the nfGO, compared to clean polystyrene (3-3.5 GPa) [15, 17], resembles the differences between normal and fibrotic tissue [28, 29]. Moreover, the negative charge that characterised GO also favourably changed the surface that imitated the negative charge of proteoglycans and hyaluronic acid, as opposed to collagen [30, 31]. The beneficial change in GO surface properties also concerned increasing their hydrophilicity by the presence of numerous oxygen groups, such as hydroxyl (-OH), carboxylic (-COOH), and epoxide (-O-) [23].
The second factor, the source of the cocktail of various proteins, was chicken liver embryo extract. The unpredictable demand of cells for signalling growth-structural factors under the influence of the modified surface (mimic ECM) determined the choice of CELE. Namely, a unique phenomenon of embryo growth and development has been used, consisting, inter alia, in creating a perfect, dynamic niche for extremely fast proliferating cells [12, 21, 32]. Most of the proteins from CELE were part of the perfect ECM/niche. CELE provided two types of laminins (LAMA1 and LAMC1), whose level is reduced in HCC [5], and also only two types of collagen (COL12A1 and COL5A1) that are detected in healthy tissue but not in cancer tissue [33]. Other important proteins include fibronectin (FN1) and vitronectin (VTN), which are responsible for the regenerative process [6, 34]. The extract also contained functional proteins responsible for ECM modification, tight cellular connections, migration, EMT, and asymmetric cell division in embryogenesis and proliferation. However, the behaviour of the CELE should be considered as a dynamically migrating and changing structure and above all one that potentially interacts in two directions. The first is direct contact and signalling to the cell, while the second is depositing the GO nanofilm and building the mimic ECM/niche structure [15, 20]. The high roughness of the GO surface enriched with π electrons can promote cell adhesion and adsorption of ECM proteins through noncovalent π-π stacking interactions [35, 36] and form the so-called protein corona [37, 38, 39].
The first line of contact between the cell and the ECM are integrins, which in response to signals from outside transmit information to the cell and are the source of the signalling cascade covering most of its functions [5, 40, 41]. Integrins, heterodimeric receptors, belong to the family of adhesive proteins and contain 18 alpha subunits and 9 beta subunits occurring in 24 alpha-beta combinations [42]. However, a high level of expression of one subunit may be sufficient to alter the expression of the entire heterodimer [43]. Nine key integrin subunits were identified in the conducted studies. Interestingly, the expression of examined integrins under the influence of both the surface and the CELE was definitely different in HepG2, C3A, and non-cancer HS-5 cells, which demonstrated the different sensitivity of various types of cells or/and tumours to mechanical and chemical signalling derived from the cell microenvironment. Thus, the mechanical characteristics of the substratum can be expressed due to the dynamics of integrin clusters in a cell-specific manner [44]. What's more, it confirms the thesis of the need for targeted therapy and the risk of obtaining a negative effect in mismatched therapy. Additionally, C3A cells cause a higher resistance to treatment than the HepG2 cell line [16]. C3A cells produce more albumin and alpha-fetoprotein than HepG2 cells and can grow in the glucose-deficient medium. Albumins and alpha-fetoprotein from C3A can adsorb onto nanomaterials, form protein corona, change the interaction nfGO-cells, and mitigate the effect of nfGO [45, 46].
Studies on integrin expression still do not allow for an unambiguous arrangement of their role and significance depending on the type of cells, type of cancer and phenotype of cancer cells, stage of cancer, and metastasis. Nevertheless, numerous studies have allowed for the creation of a map of plausible integrin heterodimers as therapeutic cancer targets [42]. Based on the analysis of the expression of selected integrins in liver tumours compared to healthy tissue, αV and β1 subunits can be indicated as the most frequently recurring in typed heterodimers [40, 43]. In our studies, the expression of β1 integrins did not change under the influence of the factors used in HepG2 cells, in contrast to C3A cells. This confirms the diverse response of different types of tumours and above all may suggest the procancerogenic activity of both nfGO and CELE for the C3A tumour line. Analysis of αV integrin expression also confirms the adverse effects of the CELE and nfGO in C3A cells. However, in HepG2 cells, the expression of integrin αV was lower under the influence of CELE, although higher under the influence of surface modification with GO nanofilm. Integrin αV is upregulated in many tumour cells, making it a promising therapeutic target. Most integrins, including αV, can bind various ligands, e.g., fibronectin and vitronectin [42]. Despite the presence of these ligands in the extract, there was no increase in expression of the αV subunit, which is considered to be a marker of liver fibrosis. In addition, the extract reduced expression of the α6 subunit, which is involved in treatment resistance, spheroid and exosome formation, and metastasis [34, 47]. An inhibitor of αV integrin is a new drug for the treatment of glioblastomas that is being verified in a phase III clinical trial [43]. However, this therapeutic method of blocking integrin antibodies is not an excellent solution and is not always effective. The reason for the ineffectiveness of such a therapy may be that integrin overexpression may be a survival signal and result from feedback (ECM - cell) in which the cell does not adhere to a degraded niche, which in turn promotes metastasis [48]. Thus, the desperate attempt of the cell to contact the ECM/niche and consequently the increase of integrin expression may be greater the more its environment is degraded. Paradoxically, despite being a kind of "striving for normality", it is proportional to the carcinogenicity and metastasis potential. Moreover, most studies do not take into account the physicochemical state of the extracellular matrix, which is the largest regulator of integrin expression and activity, especially when faced with a very large number of ligands present in the ECM and cell surface adhesion proteins that bind integrins. The presence or absence of specific ligands in the ECM appears to be crucial for integrin expression and status. Culturing HepG2 cells on nfGO reduced the expression of the laminin binding α3 integrin and fibronectin binding α5 subunits in HepG2 cancer cells [41]. Laminin and fibronectin are involved in the formation of dynamic cell binding to ECM. Furthermore, they mediate cell binding to collagen [12]. The α5β1 integrin is the most important fibronectin receptor that binds to RGD-containing peptides. It can be assumed that the GO nanofilm has become a structure that imitates laminin and fibronectin, becoming a surface that promotes HepG2 cell adhesion [15, 49]. Although it seems more likely that the protein corona, made up of proteins available in the culture medium, exposed ligand-like structures on the surface. However, by analysing the expression of α 5, overexpression of this gene was observed when CELE was added to the culture medium. This expected effect was probably the result of the increased availability of ligands (fibronectin) in the culture medium, whose source was CELE, and activation of their binding to alpha 5 integrin, however, independent of the surface. Interestingly, surface modification and simultaneous addition of CELE increased the expression of the integrin α5 subunit, which can be explained by the increased availability of ligands through binding them to the GO nanofilm. The GO nanofilm roughness was greater than the culture plastic plate, but also proteins’ noncovalent π-π stacking interactions [35] could increase the amount of bottom-related proteins. Furthermore, this interaction could be preferentially directed to the RGD-mimicking GO motif due to tryptophan, which also has π-π interactions due to the aromatic ring [35, 50]. Moreover, the use of both factors enhanced expression of the α5 subunit to the greatest extent. This integrin is involved in cell strong adhesion, and thus we paid attention to the proteins involved in cell-ECM and cell-cell adhesion mechanisms.
Mechanotransduction resulting from integrin clustering dynamics, depending on the rigidity of the substrate, is associated with FAK phosphorylation [44]. The expression of fak increased in HS-5 and HepG2 cultivated on GO with CELE supplementation. GO may reduce [41, 51] or enhance fak expression [52], which may depend on the presence of ligands, and consequently in our research, CELE was added to prevent this. Liver cells are anchor-dependent cells in which the level of fak increases in suspension culture [53, 54, 55]. The tested cells adhered tightly to the surface, so overexpression of fak resulted from increased via integrin signalling, conditioned by strong cell adhesion to nfGO + CELE, and it could be a signal of cell survival in the proadhesive but antiproliferative niche [56].
The cadherin-catenin complex carries the mechanotransduction signal between the cytoplasm of neighbouring cells [57]. High expression of E-cadherin is a characteristic feature of epithelial cells; it inhibits translocation of β-catenin into the nucleus and promotes better prognosis in cancer therapy [58]. Expression of adhesive protein genes such as e-cadherin and n-cadherin was higher under the influence of GO nanofilm and nfGO + CELE, especially in HepG2 cells, but most important was a reduction in β-catenin expression in these cells [11, 54, 59]. Our results, like Mu et al., showed that cells increase the expression of E-cadherin on nfGO, moreover, in view of the fact that the weakening of tight connections (E-cadherin expression) and an increase in flexible cell-cell contacts (N-cadherin expression) is characteristic of HCC [21]. Thus, the increase in expression of two types of intercellular junctions suggests that the experimental factors used (nfGO and CELE) did not increase the invasion of HepG2 cells [21, 59, 60]. These results are also confirmed by the observation of cell morphology, which reflects the cell-ECM and cell-cell relationship and also indicates material biocompatibility [32, 61, 62, 63].
First of all, tendencies to form smaller cell clusters and a larger number of single cells strongly associated with the GO surface were observed in both HepG2 and C3A groups. This confirms the biocompatibility of the GO plane [17] and also indicates one- and several-cell migration instead of collective migration, which is characteristic of abnormal ECM [64]. Furthermore, the roughness and elasticity of GO as a mimic ECM could advantageously imitate the natural ECM mosaic, and it was preferentially selected by all cells [15]. The effect of CELE was slightly different for HS-5 cells and liver cancer cells. This may be due to the fact that it contained more ligands and other factors needed by the liver cell rather than the connective tissue cell. Our research and that of other authors may indicate that extracts from chicken embryo [21], zebrafish [65], and amphibian oocyte [66] are excellent cocktails of factors preventing metastatic phenotype.
External mechanical-chemical stimuli, received through integrin-dependent mechanisms transmitted to the cell, create a cascade of events that determine the fate and purpose of the cell. Analysing the cell cycle, it was observed that the population in G0/G1 phase decreased only in the case of HepG2 cells. Furthermore, mcm2 and pcna gene analysis confirmed a decrease in cell population in G1 and S phases, respectively. Our research and Wang et al. showed that overexpression of the α5 subunit inhibits the proliferation and invasiveness of cancer cells [67]. The reason for this phenomenon could be the mechanism we observed via integrins and adhesion components passed on to the cell. The results obtained indicate activating Mek/Erk, PI3K/Akt, and the small GTPase Rac pathways which are involved in the synthesis of cyclins (D1 and E) and the transition of cells from the G1 phase to the S phase by integrin signalling and physical interaction of integrins with growth factors and growth factor receptors (EGFR and VEGFR). Therefore, proper cell adhesion overcame the first proliferation checkpoint [68]. In most human cancers, the G0/G1 checkpoint does not completely stop the cell cycle as opposed to G2/M [69]. In the present work, the use of two factors (nfGO + CELE) increased the population of HepG2 and C3A cells in the G2/M phase, thus preventing cells with damaged DNA from entering the M phase and allowing damage repair [16]. Ultimately, the total population of HepG2 cells in the active phase of the cell cycle was increased, in particular in G2/M phase, which confirms the increase in the ki67 gene expression. Moreover, many extract proteins (including nitrilase 2 (NIT2), DNA-binding protein 43 (TARDBP) and dynein light chain) adsorbed on nfGO or available in the medium may have been involved in arresting the cell cycle in the G2/M phase [65, 70, 71]. Our results and Jabaily et al. confirm that liver extract was a universal source of missing proteins, and in the future, it may modulate the proliferation of various cell lines including bone marrow and muscle cells [32]. The final and fundamental evidence of the loss of HepG2 cell invasiveness is a reduction of proliferation in the BrdU test of more than 40% compared to the control, which was not observed in non-cancerous cells.
All of these data indicate that graphene oxide and a cocktail of proteins that formed the dynamically changing proteome of the cell environment, migrating in the culture medium to the GO plane and the cell and therefore available multilevel (as a source of ligands and other proteins and as a modulator of the chemical characteristics of the GO surface), may have therapeutic effects on HepG2 liver cancer. ECM-therapy may have the effect of supporting traditional treatment, especially since degraded ECM characteristic for tumours becomes a pro-tumour factor and in any case hinders treatment.
{kind=link}